
The thorax was opened under ether narcosis. A. Du Bois-Raymond coil was used for the electrical stimulation. The current was not measured but was barely perceptible to the touch of the wet finger. The electrodes were first applied to the visceral pleura of the upper lobe of the left lung. The first intercostals of the left side were strongly contracted. The muscles near the second thoracic spine were also contracted on both sides. After the stimulation had been maintained for several minutes, the intercostals of the right side, the muscles of the neck, especially the scaleni and the sterno-cleido-mastoid, and nearly all of the inter-scapular muscles became somewhat contracted. The electrodes were then placed upon different areas of the visceral pleura. The intercostals normally covering the area stimulated were first contracted. Stronger stimulation initiated the contraction of the corresponding intercostals on the opposite side also. The lobes of the lung were then displaced, but not cut. The stimulation of the visceral pleura of any part of the lung was followed by the contraction of the intercostal muscles normally covering that part, and by the contraction of the spinal muscles of the same and adjoining segments. This proved that the reactions observed were not due to any local effect of the electrical current, but were true reflexes, initiated by the electrical stimulation of the sensory nerves of the pleura. Contraction of the diaphragm, the quadratus lumborum, and occasionally the abdominal muscles followed the stimulation of the lower lobes of the lungs. In the dog, the inferior lobe on the right side lies next to the diaphragm but does not touch the thoracic wall. The stimulation of this lobe initiated the reflex contraction of the diaphragm but not of the intercostals unless the stimulation was maintained for some time. The division of the phrenic prevented this last reaction. The removal of the visceral pleura in the areas stimulated rendered the reactions much more diffuse,--that is, the reflex muscular contractions were not so strong, and involved a much larger area. They were otherwise unchanged. The phenomenon probably indicates that the innervation of the pleura is more nearly exactly segmental than is the innervation of the pulmonary epithelium. This accords with circumstances of the development of these tissues. The pulmonary epithelium arises as an out growth from the pharyngeal epitheliuim, and as it grows downward it carries with it its original nerves, the vagi, and later receives its vaso-motors with its blood vessels. The pleura, on the other hand, is developed in place from the mesoblast of that region, and retains both its position and its innervation throughout life.
The section of one or both vagi above the superior cervical ganglion did not perceptibly affect the results. Section of both vagi below the superior cervical ganglia lessened the reactions in a very variable manner in different animals. After section of both vagi below the stellate ganglia, or after the extirpation of the stellate ganglia without injury to the vagi, or after the destruction of the upper part of the spinal cord, no contraction of striated muscles could be discerned upon the administration of such stimulation as we were employing. The spinal muscles were variously contracted during the application of the stimulation to the lungs. The most marked contractions were noted in the area of distribution of the second, third, fourth, and fifth spinal nerves. There is a slight appearance of segmentation in the areas of these reflex contractions. Hence, the location of the reflex muscular contractions affords some information concerning the location of areas of irritation in the lungs and pleura, especially in the early stages of disease, before the reflexes have become spread into the neighboring segments. The place of these reflexes in pathology may be referred to briefly in this connection. The irritation of the pleura by any of the causes of its disease initiates the contraction of the intercostals covering the affected area, and rest is thereby secured for the injured tissues. If the irritation is very severe, the whole thorax becomes practically immovable. Conversely, the contraction of these spinal muscles exerts an influence upon the circulation and nutrition of the injured lung which is at first curative, but becomes decidedly detrimental to recovery if the irritation be long continued.
The relations determined by these experiments were held indicative of certain possibilities in testing the nature of the somato-visceral reflexes. In the first experiments the electrodes were placed upon the skin in the area of distribution of the chosen nerve. The visceral effects thus secured were inconspicuous and inconstant. The fact that any visceral effects were secured, even though they were neither constant nor pronounced, shows that the skin may, in occasional cases, be a source of visceral malfunction through the reflex effects upon the visceral nerves. Since the skin and other superficial tissues are normally subjected to great variations in stimulation, the changes needful to produce any considerable visceral effect must be very great or else very unusual in their quality. The visceral effects following the injury of extensive skin areas are probably produced in this way. Mechanical stimulation of the sensory nerves in the deeper muscles, articular surfaces, and adjacent tissues initiated decided visceral effects. Those movements which affected the joint surfaces most urgently were most effectual in producing vascular changes. The thorax remaining intact, and the abdomen being opened, the color of the lungs was noted through the central tendon of the diaphragm. In dogs, this tendon is usually very large and thin, and the changes in the color of the lungs can be seen quite easily in a good light. In cats, the central tendon is smaller and thicker. Hence the cat is not a proper subject for this experiment. In dogs, mechanical stimulation of the deeper tissues near the fourth and fifth thoracic spines was followed, after a latent period of from one to three minutes, by a lightening in the color of the lungs. After a few minutes rest they began to redden again, becoming somewhat deeper in color than they were before the stimulation. The stimulation was again given in the same area, when the lungs again became lighter, only to redden again under rest. There seemed to be no limit to the number of times the reaction could be repeated, provided the anesthesia were not too profound, and provided sufficient time were given for the return to the normal condition between the periods of stimulation. It is evident that the changes observed were due to changes in the caliber of the pulmonary blood vessels. Experiments performed in the same manner, except that the stimulation was applied to other spinal areas, or to the trunk of the vagus, were not successful. No stimulation of the vagus seemed to exercise any effect upon the circulation through the lungs except those evidently attributable to cardiac effects. Certain effects were produced upon the bronchial musculature by stimulation of the vagus which required further investigation. Again, with the thorax intact and the abdomen opened, the fourth and fifth thoracic spines were held firmly in such a manner as to force them slightly out of their normal alignment, without actually dislocating the vertebrae. Thus the artificial lesion was produced. After a latent period somewhat longer than before, the lungs became redder than normal. This appearance persisted as long as the lesion was maintained. If the lesion were removed quickly, the normal color slowly returned. The longer the lesion was maintained, the longer the time required for recovery. Recovery from the second experiment upon the same animal was very slow, and the animal often died before recovery from the third experiment, even though the lesion had been maintained each time only long enough for the effects to become manifest. In the next series the thorax was opened. The same effects followed the same manipulations. Stimulation of the tissues near the second and third thoracic spines seemed to produce greater effects upon the circulation in the upper lobes of the lungs than did the stimulation of the tissues near the fourth and fifth spines. No actual measurements were possible under the conditions of our experiments, but differences in the color changes seem to warrant the statement just made. It is apparent that the somato-visceral reflexes are much more diffused and irregular than the corresponding viscero-somatic reflexes. Stimulation of the tissues near the second and third thoracic spines affected many other tissues also, and for this reason it seems probable that the centers most effective for modifying the circulation through the lungs are found near the origin of the fourth and fifth thoracic nerves. In order to eliminate cardiac effects, the cardiac nerves were all divided The reactions described above did not vary after the section, except that the animal did not live so well under anesthesia, and the experiments could not be so often repeated.
The following experiments upon human beings can be repeated quite easily. The subject was permitted to rest quietly until the blood pressure remained constant. The blood pressure was then estimated by means of the sphygmomanometer. Stimulation of the tissues near the origin of the fourth and fifth thoracic nerves was followed by a rise in blood pressure, such as would be caused by the contraction of the blood vessels in any marked area of the circulatory system. In the light of the experiments upon animals just described, it appears that this increase in blood pressure is due to the decrease in the size of the pulmonary vessels. Deep, steady pressure at the sides of the spines, or the maintenance of an artificial lesion in the same area, is followed by a decrease of blood pressure, such as would follow a dilatation of the vessels in any large area of the vascular system. This effect is, for the same reasons, held to be due to the lessened action of the pulmonary vaso-constrictors. Figure 4 and figure 5 show the effects of these manipulations upon the respiratory curves. The subjects were permitted to remain quiet upon a treating table until no further change in pulse or respiration could be detected. The normal curve was then taken, then the manipulations administered, then a second tracing was taken, with the same apparatus similarly placed. These curves were taken many times with many different persons as subjects, and the results were always alike, in the main, though they differed considerably in detail. In many instances, the curves varied in answer to the manipulations very much more than do those which were used in the cuts. These were chosen as representative of the average changes produced by the manipulations described. The respiratory curves were taken with Mareys tambour. This consists of two pans connected by rubber tubes. Both pans were covered with thin sheet rubber. One pan was bound over the apex beat of the heart, the other supported a needle which played upon a smoked paper around the revolving drum of a kymograph. The pressure of the respiratory movements and of the heart beat was transmitted through the fluid in the first pan, then through the tube to the second pan, where the needle was affected, and the needle wrote the record upon the smoked paper. The paper was then varnished and the records preserved. The large waves are those produced by the respiratory movements, and the smaller waves superimposed upon the respiratory curves are made by the heart beat. 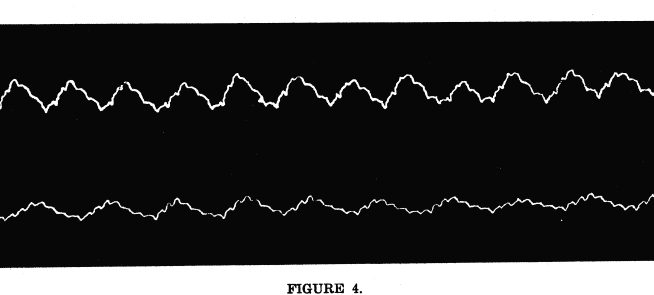 Upper tracing, normal, resting, respiratory curve. Blood pressure, 110 m.m. of mercury. The smaller waves were made by the hearts beat. Lower tracing, respiratory curve after the maintenance of an artificial lesion of the fourth thoracic vertebra for three minutes. Blood pressure, 99 m.m. of mercury. The lesion causes slight dilatation of the pulmonary vessels, increased oxygenation of the blood, and decreased activity of the respiratory center. The decreased blood pressure is due to the dilatation of the pulmonary vessels.  Lower tracing, resting, normal respiratory curve. Blood pressure, 116 m.m. of mercury The smaller waves were made by the hearts beat. Upper tracing, respiratory curve after stimulation of the tissues near the fourth thoracic spine. Blood pressure, 121 m.m. of mercury. The stimulation caused constriction of the pulmonary vessels, decreased oxygenation of the blood, and increased activity of the respiratory center. It was noted in the observations upon anesthetized animals that the reflex effects upon the heart disappeared much more quickly under the anesthesia than did the vaso-motor reflexes That is, under anesthesia the cardiac nerve centers first lose their power of replying to sensory impulses, while the irritability of the vaso-contrictor centers persists under more pronounced anesthesia. This is true also of the same nerve centers during the process of dying under certain other poisons. The experiments upon human beings were, of course, carried on without anesthesia. The cardiac centers were therefore, normally active. When the human subject slept, however, the cardiac reflexes were less conspicuous than the vaso-motor reflexes, just as in the case of the anesthetized animals. It seems that sleep and anesthesia alike differ from the waking condition in this particular case, as well as in others which are more familiar. In these experiments upon human beings, the effects described in this chapter were constant only as described if the subject were in fairly normal condition. If the inter-scapular muscles were contracted or if the manipulation was painful the effects varied in some instances. Any manipulation, whether stimulating movements or steady pressure, seemed to effect the relaxation of the muscles, and a return to the normal condition. These effects are explained as follows: During the period of the abnormal contraction of the inter-scapular muscles, however produced, the reflex effect upon the pulmonary vessels was such as was produced by the steady pressure of the experimenting fingers. The vessels were dilated in the same manner as was seen in the vessels of the animals subjected to the same experiment. Any manipulation of these muscles resulted in their relaxation, and in the return of the whole circle of sensory, vaso-motor and associated neurons and the vascular musculature to the normal condition. It seems that contracted muscles or bony lesions in the area of the lung center or its immediate neighborhood exert an inhibiting influence upon the vaso-motors of the lungs, leading to their dilatation, and to low systemic arterial pressure. The relaxation of these muscles or the removal of the lesions, however secured, removes the source of the abnormal sensory impulses. Then the circulation becomes normal as soon as the vascular walls and the neurons affecting them recover from the effects of the abnormal influences.
The Pulmonary Vaso-Motors, Dr. Pearl A. Bliss, A. O. A. Journal, August, 1907. |