|
The blood vessels of the cranial structures receive their nerve impulses from a very complex arrangement of nerve centers in the floor of the fourth ventricle and the acqueduct of Sylvius, and in the spinal cord of the upper thoracic region. These centers are not described very clearly in the ordinary text books of physiology, and therefore a brief and simplified description of their relations will be given here for the sake of making the significance of the experimental work intelligible to those whose study of the relations of the cranial sympathetics has not been unusually thorough. It is not possible, within the limits of this volume, to give an exhaustive description of these relations. A description of the manner in which the vessels of the nasal membrane receive their vaso-motor impulses will serve as an illustration of the innervation of the vessels of all the cranial structures. The coordinating centers for the vascular innervation of these membranes, as for all other structures of the body which receive vaso-motor nerves, are affected by afferent impulses from the structures whose circulation they control, by impulses from other centers in structural and functional relationship with these, and by impulses from certain ganglia grouped around the base of the brain. The sensory impulses carried over the olfactory nerve are of value chiefly because of the impulses which they send to the cerebral cortex, and the information thus given in consciousness. There is reason to believe that olfactory impulses do initiate some efferent impulses to the nasal mucous membrane, but these are not yet well understood. Their further discussion is of no immediate interest in this connection. The
fifth cranial nerve is the nerve of common sensation to the cranial structures.
Since the nerves of common sensation are the ones which most efficiently
carry impulses aroused by external changes to the vaso-motor centers, the
fifth is the nerve chiefly concerned in the afferent side of the path of
the nasal vaso-motor reflexes. Stimulation of the sensory endings
of the nasal branch of the ophthalmic division of the fifth nerve by irritating
gases or dust reaching the nasal mucous membrane causes an increased secretion
of the nasal mucous glands and a congestion of the nasal membranes.
The pathway traversed by the impulses concerned in this familiar reaction
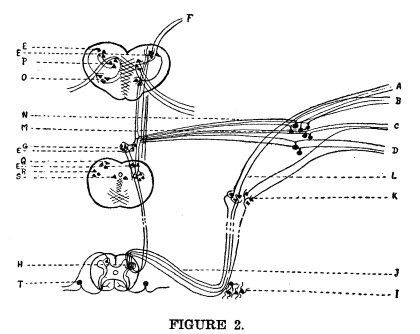
is shown in the following diagram.
Some of the axons from this nucleus carry impulses by way of the fillet, F, to the optic thalamus, whence they are sent to the cerebral cortex, where consciousness is affected; other axons carry impulses to the vaso-motor center, G, in the lower triangle of the floor of the fourth ventricle. The exact location of the vaso-motor center is not known, but since the axons and collaterals from the nucleus of the fifth enter practically all areas of the gray matter at all levels within the bounds which are known to include the vaso-motor center, the relations are unquestionably as illustrated. From the vaso-motor center impulses are carried by axons which pass downward through the lateral ground bundles of the spinal cord to the upper thoracic region. Here the axons which carry the impulses concerned in the regulation of the size of the vessels of the nasal mucous membrane plunge into the gray matter of the cord, and terminate by forming synapses with the neurons of the lateral horns in the second and third thoracic segment, H. The neurons of the lateral horns send their axons by way of the white rami communicantes, J, into the sympathetic system. The axons in which we are now interested do not enter into relations with the sympathetic ganglion cells until the superior cervical ganglion, K, is reached. The axons of the cells of the lateral horns which carry impulses for the control of the cranial sympathetics retain their medullary sheaths through the upper dorsal and cervical region until they reach the superior cervical ganglion. Here they break into fine fibrillae, which form networks around the bodies of the sympathetic neurons. The axons of the sympathetic neurons are not medullated. Those concerned in nasal vaso-motor impulses pass to the nasal branch of the ophthalmic division of the fifth nerve, and with it reach the vessels of the membranes whose sensory nerves were first stimulated. It seems from the appearance of slides prepared in these laboratories that several cells of the sympathetic ganglia may be affected by one axon from the white ramus, and that more than one axon from the white ramus may affect a single sympathetic cell. This structural relation renders possible the very diffuse manner in which visceral reflexes are produced. (Figure 3). It must be plainly understood that the sympathetic system is not in any sense a complete system, but that it is merely a way station for the transmission and diffusion of impulses from the viscero-motor centers in the spinal cord, medulla, pons, and mid-brain. It must be understood, also, that none of these centers has any volition at all, but that every efferent nerve impulse is the inevitable result of the structural and functional relationships of the centers concerned in the reaction.
The dendrites of the sympathetic neurons and the fibrillae from the axons from the white ramus make a perfect rete mirabile. It is probably because of this marvelous complexity and the extreme tenuity of the fibrillae that the relationships here illustrated have not been described before.
In the first series of experiments, the electrodes were placed upon the nasal mucous membrane of animals under anesthesia. The muscles near the third thoracic vertebrae were at once strongly contracted. The latent period was very short in all reactions involving the cranial structures. The cervical muscles were somewhat contracted in more than half of the animals subjected to the tests. The electrodes were then placed upon the conjunctivae. The muscles near the second vertebra were then contracted. There were also slight and inconstant contractions of the cervical muscles. The electrodes were placed upon the eyeball. The muscular contractions were sometimes noted near the second thoracic vertebra, but the reaction was not constant. The cervical muscles were scarcely contracted at all. The electrodes were placed upon the outer surface of the eyelids. The facial muscles were contracted very quickly and forcibly, but no contraction of the muscles of the upper dorsal region was noted. The electrodes were placed upon the tongue, the inner surface of the cheeks, and the roof of the mouth. The facial muscles were quickly contracted, and reflex muscular contractions were noted near the third thoracic spine. The electrodes were placed upon the tonsils. The reflex muscular contractions appeared near the third and fourth thoracic spines. The electrodes were placed within the pharynx. Muscular contractions appeared near the third thoracic spine. The throat was exposed to view, and the electrodes placed upon the vocal cords. The cervical muscles were very strongly contracted, as were also the spinal muscles of the fourth and fifth thoracic segments. The first and second intercostals were also strongly contracted. The trachea was opened and the electrodes placed upon its inner surface. The scalene and the sterno-cleido-mastoids were very strongly contracted. The first and second intercostals , and nearly all of the inter-scapular muscles were also contracted. The electrodes were placed upon the thyroid glands, and the reflex muscular contractions appeared near the second and third thoracic spines. The contractions initiated by the stimulation of the thyroids were not very strong, but were constant. The deeper cervical muscles were always involved in the reflex action caused by the stimulation of the thyroids. The skull was opened and the electrodes placed upon various portions of the meninges. The facial muscles were contracted in all instances, and the cervical muscles were often contracted also. The upper dorsal muscles were not often contracted unless the brain itself were stimulated. In this case, the effects of the stimulation depended upon the area of the cerebral cortex affected.
The superior cervical ganglion was exposed to view, and the electrodes placed upon it. The pupils became greatly dilated, the conjunctivae because lighter in color, and the mucous membranes of the nose and throat were also lightened. The superior root of the superior cervical ganglion was cut. The direct stimulation of the ganglion produced no effects. The sympathetic chain was cut below the superior cervical ganglion. Direct stimulation of the ganglion produced the same effects as before. Therefore the pressure of abnormally contracted cervical muscles upon the superior cervical ganglion may exert an evil effect upon the cranial structures which receive their innervation by way of these ganglia.
After section of the sympathetic chain, stimulation of the cranial structures did not produce any effect upon their blood vessels, except such as might be referred to the direct influence of the electricity upon the vessels walls. Therefore, the impulses concerned in this reflex action pass upward through the cervical sympathetic chain.
The Gasserion ganglion was exposed to view. The ganglion was stimulated directly. The upper thoracic muscles were very strongly contracted, and the blood vessels in the area of the distribution of the fifth nerve were immediately and strongly contracted. Some of the sympathetic fibers are carried by way of the fifth nerve. In order to exclude the effect of the direct stimulation of these fibers, the fifth nerve was cut, and the central end was stimulated by the electrodes. The muscles of the upper thoracic region were contracted, as before. The vessels in the area of distribution of the fifth nerve were contracted after latent period of a minute or so.
The Gasserion ganglion could be extirpated in some animals without perceptible injury to the sympathetic fibers from the superior cervical ganglion. This must have been due to some peculiarity of distribution of the sympathetic fibers. The operation was considered to be successful when the direct stimulation of the upper thoracic sympathetic cord produced effects upon the pupils and the blood vessels of the mucous membranes of cranial structures. In some animals, the sympathetic fibers are so closely connected with the fifth cranial nerve that the extirpation of the ganglion prevents all passage of nerve impulses from the cervical or upper thoracic ganglia to the cranial structures. The tests made upon the animals in whom the extirpation of the Gasserion ganglion was possible without injury to the sympathetic fibers will be considered at this time. After this operation, the stimulation of the cranial structures did not produce any perceptible effects upon the vessels of the tissues stimulated, or their neighbors, except such effects as might be attributed to the direct effects of the electricity upon the vessel walls. The stimulation of the fifth nerve did not produce any contraction of the muscles in the upper thoracic region after this operation. The stimulation of the central end of the injured nerve caused strong muscular contractions in the upper thoracic region, and also constriction of the vessels in the area of distribution of the fifth. Direct stimulation of the superior cervical ganglion produced effects identical with those produced before the mutilation. The spinal cord was cut above and below the superior cervical ganglion. This cut was made from behind, and the sympathetic chain was uninjured. The effects noted after both operations were the same, and can be described as one. The stimulation of any cranial structure failed to cause reflex contraction of the muscles in the upper dorsal or the cervical region. Stimulation of the cranial structures did not produce any vascular changes except those which might be referred to the direct effects of the electricity upon the vessel walls. Direct stimulation of the superior ganglion produced the effects noted before mutilation. Therefore, the cervical portion of the spinal cord is an essential element of the reflex arc by way of which sensory impulses from the cranial structures are able to effect the condition of the upper dorsal muscles, and also in the path by which these impulses are able to affect the size of the blood vessels of the cranial structures themselves.
The relations demonstrated in the series of experiments just described were held to be suggestive of certain possibilities of somato-visceral reflexes. Mechanical stimulation of the tissues near the second thoracic spine was followed by a contraction of the blood vessels of the cranial mucous membranes and the conjunctivae, by a dilatation of the pupils, and an increased secretion of saliva. These effects were practically invariable. The artificial lesion affecting the tissues near the second thoracic vertebra produced a dilatation of the blood vessels which was most marked in the conjunctivae, but which affected the other cranial membranes to some extent. Stimulation of the tissues near the third thoracic vertebra was followed by a constriction of the vessels of the nasal mucous membranes and the pharynx. The effects upon the conjunctivae were less conspicuous. The artificial lesion affecting this vertebra caused a slight rather general congestion of the cranial membranes. Stimulation of the cervical tissues did not produce any effects upon the cranial structures unless the stimulation was so pronounced that the cervical sympathetic chain or ganglia were directly affected. Stimulation of the muscles in the cervical region affected the sympathetic structures very easily, hence, the existence of abnormally contracted muscles in the cervical region is a menace to the normal activity of the cranial sympathetics. The artificial lesion affecting the cervical vertebrae caused contractions of the cervical muscles rather readily. In the lower spinal areas the artificial lesion affected the spinal muscles only slightly, within the time occupied by our experiments, but the cervical muscles become contracted within a few minutes after the artificial lesion was produced. The latent period for all cervical and cranial structures is comparatively very short. The
superior cervical ganglion was subjected to mechanical stimulation by the
manipulation of the tissues over it. In animals, this maneuver was
followed by dilatation of the pupils, and by a contraction of the cranial
vessels, which was soon followed, if the stimulation was continued, by
a dilatation which was rather persistent.
After the extirpation of the Gasserion ganglion without the injury of the sympathetic nerves, the mechanical stimulation of the tissues near the second and third thoracic vertebrae caused the same vaso-constriction and pupilo-dilation as was observed in the animal before mutilation. After the destruction of the cervical portion of the sympathetic chain, and after the extirpaton of the Gasserion ganglion in most animals, the mechanical stimulation of the tissues in the upper dorsal region did not produce any perceptible effects.
After the destruction of the cervical portion of the spinal cord, stimulating movements in the upper dorsal region produced the same effects upon the blood vessels and the pupils as did the same movements before mutilation. In some instances, the reactions were increased after the destruction of the cord, as was to be expected. It was not possible to make quantitative tests of these reactions, under the conditions of our experiments.
A series of experiments were performed upon human beings. Irritation of the skin over the back of the neck caused temporary dilatation of the pupils. The most conspicuous results were secured when the irritation (a brush was used) was applied to the skin over the seventh cervical to the second thoracic spines. Mechanical stimulation of the tissues near the second and third thoracic spines caused dilatation of the pupils and contraction of the vessels of the cranial mucous membranes. Inhibition, or the maintenance of an artificial lesion, caused dilatation of the vessels of the nasal mucous membranes and of the conjunctivae. The eyeball was also somewhat congested The pupils were dilated in this case also. The superior cervical ganglion is subject to direct stimulation in most persons. The fingers were firmly placed over the ganglion, and the deep vibrating movements given. In some cases, the electrodes were firmly pressed over the ganglia, one on either side. The effects of this procedure were very pronounced. The blood vessels of the conjunctivae, the mucous membranes of the nose and mouth, were all first lightened, then, if the stimulation were continued, reddened and congested with blood of rather a purple color, as if it were flowing more slowly than its custom. The flow of saliva was increased. The pupils attained very great size, if the stimulation were continued for a few minutes The upper lids were drawn upward and the eyeball protrudes The tears were increased, and often overflowed. All of these effects are due to the stimulation of the superior cervical ganglion. Since the effects are so pronounced when this stimulation is given for only a few minutes, the effects upon the cranial vascular system of abnormally contracted muscles pressing upon the sympathetic ganglia can certainly not be disregarded with impunity. It is extremely difficult to secure adequate inhibition of the anterior cervical structures, because of the propinquity of the pulsating carotid arteries. Stimulation of the tissues over the cervical vertebrae in persons, as in animals, caused contractions of the cervical muscles, but no effects upon the cranial structures were discerned, except such as could be referred to the effects of the pressure of these contracted muscles upon the sympathetic trunk and ganglia.
In no case were we able to affect mental conditions by manipulations planned to stimulate or inhibit the action of vaso-motors to the brain. Changes produced in the size of the vessels of the cranial structures sometimes caused headache and discomfort, but not ever any consciousness of sleepiness or of increased mental vigor, such as accompanied the various manipulations affecting blood pressure which were made upon the abdominal viscera These experiments do not deny the existence of vaso-motor nerves to the brain, but they do not demonstrate them. So far as our present knowledge goes, we must consider that the circulation through the brain is most easily affected through changes in the general blood pressure and that this is most easily controlled by affecting the size of the blood vessels, in the splanchnic area, and in some cases, by affecting the efficiency of the heart beat.
The superficial or osteopathic centers for the control of the cranial structures may thus be located as follows: For the eyes, the level of the first and second thoracic vertebrae. Lesions of the vertebrae or muscles higher and lower than these may affect the eye tissues also. The eyes may be affected by direct manipulation of the superior cervical ganglion, and lesions affecting this ganglion may affect any of those structures of the orbit which receive innervation by way of the sympathetic system. For the nasal mucous membrane, the second and third thoracic vertebrae. Lesions higher and lower than these, and lesions affecting the superior cervical ganglion may also affect the circulation of the nasal membranes. The relation of upper thoracic and cervical lesions with adenoids is very significant. For the pharyngeal structures, the third and fourth thoracic vertebrae. Here, as in the other cases, lesions of the vertebrae above and below those mentioned may affect these structures. Lesions affecting the middle and superior cervical ganglia directly may exert an evil influence upon the circulation and metabolism of these structures. The centers for the trachea and larynx are not to be at the level of the third thoracic. Lesions affecting the nerves of this and neighboring spinal segments, as well as lesions affecting the middle and superior cervical ganglia, may affect the thyroids through their nerves Other lesions affecting the thyroids by direct impingement upon their vascular and lymphatic drainage were not subjected to experiment. The centers for the trachea and larynx are not to be distinguished from those for the upper lobes of the lungs.
Olfactory Motor Paths, in Morats Physiology of the Nervous System, p. 643, Edition of 1906. Vaso-Motor Nerves and Reflexes, in any text book of Physiology. |