
|
The word erythrocyte means red cell, and these structures are often so called. By transmitted light, in thin layers, they are not red but a pale yellowish green tint. By reflected light, and in great masses, they appear a brilliant scarlet color, varying in tint according to several physical condition. Variations in the color of the red cells, and hence
of the blood itself, are due to modifications in the chemical relations
of the hemoglobin. Venous blood contains a greater amount of the purplish
hemoglobin; arterial blood contains a greater amount of the more brilliant
scarlet oxyhemoglobin. The peculiar cherry-like tint of the blood of persons
who are suffering from severe acute carbon-monoxide poisoning is easily
recognizable. A very much less pronounced cherry-like tint is present in
the blood of persons who suffer from mild, chronic carbon-monoxide poisoning.
Adult human erythrocytes, or red blood cells, occupy a peculiar place in biology. They have no nuclei during their most important functional activity and they are unable to take up nutrient materials from their surrounding plasma or to give off katabolites while they are most active in carrying oxygen. They have neither motility nor power of reproduction. These cells consist only of a very delicate stroma which holds the hemoglobin in its meshes. Although the erythrocytes seem almost structureless they display some characteristics of living cells. They take up and give off oxygen with greater facility than can be explained on the supposition of a purely physical basis for their functional activities. Solutions of hemoglobin have not yet been made to perform the reactions with such facility as do the normal erythrocytes. The manner in which they may be affected by pathological conditions greatly resembles the manner in which these, or similar, abnormal states act upon other cells. It is true that all of these facts might be explained by the extreme delicacy of the stroma, the finely balanced relations of the osmotic tensions of cell and plasma, and other physical conditions. These considerations apply equally well to all forms of living structures. The erythrocyte is peculiar in that it performs its most useful function,--that of carrying oxygen,--after it has become cytologically, a senile cell. Having lost its nucleus, its power of taking up nutrition and its ability to reproduce itself, it has left only its special activity, due to its hemoglobin content, of taking up oxygen when the oxygen tension of its environment reaches a certain pressure, and giving off oxygen when the oxygen tension of its environment diminishes below a certain point. In some lower animals the oxygen-carrying pigments
are carried in solution in a circulating fluid. Red blood cells are present
in the blood of all vertebrates except amphioxus, but not in the blood
of invertebrates. Invertebrate blood cells occasionally show a trace of
hemoglobin or of other oxygen-carrying pigments, but these are too few
and the amount of pigment too scanty to give any tint to the blood or the
other fluids of the invertebrate body, or of the amphioxus. The practice
of carrying the oxygen-carrying pigment within circulating cells increases
the efficiency of that pigment very greatly.
The form of erythrocytes has been rather exhaustively studied. When carefully examined upon a warm stage, under as nearly normal conditions as possible, they are seen to be shaped like a shallow bowl, with a very thick rim and a very thin bottom. The smear as ordinarily taken shows erythrocytes of the biconcave form usually described. At various times a cell wall or a peripheral limiting membrane has been described: this is a delicate condensation of the stroma. (Plate I) When moving blood is watched under the microscope, especially on the warm stage, the erythrocytes are seen to change in shape remarkably. They elongate to a surprising extent, often attaining a length of more than twice their ordinary diameter in passing through very narrow places, and regaining their normal form immediately upon reaching a widened space upon the slide. This elasticity is due to the lipoid structure of the stroma and it facilitates diapedesis through the walls of the blood vessels and within the tissue spaces of the body. The form of red cell found in human blood is the most efficient form known, so far as the transmission of oxygen is concerned. The factors which maintain this peculiar structure are not well understood. It is known that abnormal environmental conditions may so affect the red blood cell as to cause it to change shape very quickly. A typical normal adult human erythrocyte has a diameter of about eight microns, with variations of about one micron in each direction. The constant variations in the reaction of the blood and its carbon dioxide content cause variations in the size and the form of the red cells. The thickness of the heavy rim is about 1.7 microns while the central area has a thickness of less than one micron. The surface area has been estimated by several observers and the figures differ somewhat, as is to be expected since the form and size of the red cells differ according to so many conditions. The figures given for the surface area range between 98 square microns and 128 square microns. The volume of an average normal adult erythrocyte is somewhat less than 100 cubic microns. Welcker estimated the volume as 72 cubic microns, Ponder as 110 and Wintrobe as 70 cubic microns. The surface area of the blood cells provide about
4,000 square meters for an average human body and of this about 81 square
meters of surface pass through the lungs each second. Evans computed 3,500
square meters and Bailey 4,500 square meters of erythrocyte surface for
an average human adult blood volume.
The red cells vary slightly in different individuals, with no apparent relation to physiological conditions. Individuals who are developmentally imperfect have greater variation in the size and form of their cells during health than do persons who have normal structure of body. Conspicuous abnormalities of structure are rare in healthy persons. Individuals with oval blood cells have been reported in several instances. Mrs. L., patient in the obstetrical clinic of The Pacific College of Osteopathy had oval cells almost exclusively. Only occasionally was it possible to find a circular erythrocyte in her blood. The long axis was about one and one half times the short axis; the relation varied slightly. No nucleated forms were present. Seven blood examinations were made during four months, giving identical findings. She was in good health, passed through normal pregnancy and labor, bore a normal child and regained her strength as rapidly as is usual. Her hemoglobin varied slightly, from 82% to 88%, but she never showed any evidences of anemia. Her family history was excellent and the family records were complete for several generations, both direct and collateral. With the good health common to nearly all members of the family there was no history of blood examinations having been made. The examinations were made twenty-two years before this history is written, and she is still in excellent health. In one monkey (rhesus) in the Chicago laboratory
of The A. T. Still Research Institute oval erythrocytes were found present
in the peripheral blood in considerable numbers. This monkey was in as
good health as monkeys ever are under artificial conditions. The cells
resembled the cells of the patient whose history has just been given.
Races living in the tropical zone have red cells about 0.5 micron smaller than races living in temperate zones and they have slightly lower counts of red cells. Races living in very cold climates have red cells of about the same size as those of temperate zones, but their red cell count is somewhat higher. The red cells vary in size and in form during the day, following variations in the reaction of the blood. The amount of hemoglobin in each cell does not vary, however. During activity and the diminished alkalinity of moderate fatigue, the red cells increase in size. Forced breathing with the associated slight increase in alkalinity causes the red cells to shrink slightly. Increase in the carbon dioxide content of the warm stage specimen causes cells previously shrunken to return to their normal size and form. Diluting fluids which are slightly more alkaline than normal blood cause the red cells to crenate more rapidly than normal, while diluting fluids less alkaline than the blood cause them to become more globular, then to swell. At very low carbon dioxide tension this gas is carried almost exclusively by the blood plasma, and in such blood the cells crenate rapidly. With higher carbon dioxide tension the gas is carried by the plasma and the red cells in about equal amounts, and under such conditions the red blood cells do not crenate so rapidly. Resistance of red cells to lowered osmotic tension of the plasma, caused by adding water, is increased by raising the carbon dioxide tension. The form of the red cells depends to some extent upon carbon dioxide tension and this is, no doubt, the reason for so many different descriptions of these cells. Experimentally, substances which dissolve the lipoids
of the stroma cause the cells to assume forms which are almost or quite
spherical. The presence of bile acids in the plasma of the circulating
blood is frequently associated with the presence of spherical or spheroidal
red blood cells.
The erythroytes of newly born healthy infants range from 3.3 to 10.0 microns in diameter, with an average of 8.6 microns. During childhood the cells become more and more nearly equal in size until the adult type is reached at the age of about fourteen years. Fetal blood shows even more marked variations in size than does infants blood. During childhood the red blood cells show extravagant variations in size and form upon relatively slight provocation, so that even mild cases of secondary anemia may occasionally cause the red blood cells to show the anisocytosis and megalocytosis characteristic of pernicious anemia if they should appear in adult blood. It is necessary to keep this characteristic of immature blood in mind, else erroneous diagnoses of pernicious and other very severe anemias may be made for children with secondary anemias. In old age the red cells become somewhat smaller.
This seems to be due to the increased alkalinity of senile blood.
Conditions which interfere with the nutrition of the red bone marrow cause the appearance of abnormal forms of red blood cells. These conditions include bony lesions, which interfere with the innervation of the red bone marrow and the circulation of the blood through it; disturbances in the nutrition of the entire body, as by starvation or by disease of the digestion tract; diseases which cause severe toxemia and other conditions which cause secondary anemia. The abnormal forms found among the red cells in anemia are sometimes due to abnormal development of these cells, but are often due to their abnormal fragility. In certain forms of anemia the cells are so fragile that it is extremely difficult to prepare them for examination without injuring them, yet, if the cells are very carefully prepared, nearly all of them are found to be of normal size and form. In the primary anemias the cells are often more stable
in structure than they usually are in the blood of normal persons, and
in certain forms of secondary anemia the resistance of the red cells to
abnormal environmental conditions may be considerably increased.
The red cells of normal adult human blood present only the single structure described,--a fine and very delicate stroma of lipoids holding in its meshes the hemoglobin. The delicacy of this stroma permits its easy modification by various fixing agents and stains, so that many and varied intracellular structures have been described. Immature and abnormal forms present rather complicated structures and these have been variously described by different observers. Living red cells do not take stains. Fixed and dead adult red cells are acidophilic. Immature red cells and abnormal forms are often feebly basophilic. Polychromatophilic or basophilic cells take basic stains. This term is not properly applied to basophilic reticulation. Basophilic red cells are usually larger than normal; have less marked concavity, and are often poikilocytes. Very young red cells are strongly basophilic and usually show nuclear remnants. Degenerating red cells may become basophilic, as in severe anemias, especially when these occur in old people. Golgi described a reticulo-fibrillar apparatus which he found in red cells stained by two methods, one using mercuric chloride with potassium bichromate, the other using gold chloride with picric and osmic acids. These methods produce a fibrillar or reticular structure. In our laboratories these appear very distinctly to be artifacts, produced by the action of these agents on the stroma of the red cells. Petrone used a lead-impregnation method and produced an endoglobular body which also seems, in our slides, to be an artefact. The differentiated inner body of Lowit has a somewhat fibrillar structure; it is found only in immature forms and is supposed to be a remnant of the nucleus. Nucleoids are remnants of the nucleus, in cells not yet quite mature. Morris granules are single, rather large sharply circumscribed basophilic granules lying near the center of the cell, giving with all stains the reaction characteristic of nulei; they are very probably nuclear remnants. These are normally present in embryonic blood and in adult blood during rapid hematopoiesis. Vaughan nuclear remnants are basophilic masses which
lie near the center of the cell; they are probably nuclear remnants and
are increased in conditions associated with rapid hematopoiesis. They may
occasionally be seen in normal blood.
Several kinds of granulations appear in the red cells. In malaria fragments of the chromatin of the parasites appear as granules; in old, atypical cases of malaria these may present considerable difficulty in diagnosis. Fragments of disintegrating nuclei are found in the peripheral blood during rapid blood regeneration. Various precipitation forms of abnormal erythrocyte protoplasm are occasionally found. Under abnormal conditions erythrocyte protoplasm presents atypical structures which may be granular. Platelets and other plasma constituents often adhere to erythrocytes and appear to lie within them. The various granules, artifacts and structural peculiarities thus produced within the erythrocyte have received many names; some of these names indicate definite conditions. In other cases different names are applied to the same structure; perhaps with a different appearance due to differences in staining technique. It is not now possible to clear up the problems presented by these endoglobular structures; it may be convenient to review very briefly the terms most commonly applied to them, though it is not possible to explain their presence. Maraglianos endoglobular degeneration consists of areas of definitely basophilic protoplasm occasionally found within the red cell, which itself is normally basophilic during its youth. The persistence of this immature structure occurs in many anemias. Cabots rings are sometimes circular, sometimes oval, sometimes long, slender, almost band-like in form. Cabot thought them nuclear remnants. They are especially abundant in pernicious anemia. Grawits basophilic degeneration is probably not a true degeneration but an indication of immaturity. Cells showing this granulation are often called stipple cells. There are fine, dust-like or granular particles of basophilic material within the cells which are present in the blood in nearly all cases in which blood regeneration is proceeding rapidly, especially in pernicious anemia and in lead poisoning. They occur normally in the red bone marrow. Ehrlichs hemoglobinemic degeneration seems to consist
of denser particles of hemoglobin, probably of abnormal structure, within
the cell, itself much paler than normal. The conditions resemble those
which might be expected to occur if the hemoglobin were to be collected
together in masses within the stroma of the cell.
When red cells are placed in a fluid of increased or decreased alkalinity or of increased or decreased osmotic tension they undergo several changes. In hypotonic fluids with alkalinity slightly less or equal to that of the blood, the cells swell slightly and become more nearly or quite spherical. The hemoglobin diffuses out from the cells leaving only a ghost or colorless remnant composed chiefly of the lipoids which make up the erythrocyte stroma. This phenomenon is called laking. In hypertonic fluids with alkalinity equal to or
slightly greater than that of the blood, the cells give off water, shrink
and become more nearly spherical. The surface is thrown into rounded or
sharp prolongations somewhat resembling a burr; this phenomenon is called
crenation. These prolongations may even become pinched off from the mass
of the cell somewhat resembling a strong of beads. Laking occurs under
such circumstances much less rapidly than is the case when the cells swell
in hypotonic solutions.
When the blood is placed upon a slide at 100 degrees F., which is about the normal temperature of blood, and the slide is covered and sealed to prevent evaporation, the red cells retain their normal size and contour for a long time. If the slide is too cool, at about 97 degrees F., the cells shrink slightly and become more nearly spherical. If the slide is too warm, at about 105 degrees F., the cells become fragmented. Most commonly the cells become somewhat more nearly spherical, then a constriction appears near the equator. This increases in depth and finally the cell is divided into two parts. The constriction may appear near one pole of the cell in which case two unequal masses result. Rarely the cell breaks into three or four masses at about the same time. The cells assume various bizarre forms; commas, kites, dumb-bells and many other strange shapes occur. Certain abnormal forms of red cells appear during the course of anemias and these may be briefly described. (Plates I, VII, VIII) Normocytes are red cells whose diameter varies only within the limits normal to the age of the patient. This age question is of importance, because in children the normal variation is greater than in adults. Microcytes are red cells less than six microns in diameter; they may be as small as three microns in diameter, and even smaller forms are sometimes found in stained dried smears. Megalocytes and macrocytes are large red cells, and the terms are used interchangeably by many authors. Certain authors have restricted the term megalocytes to cells eleven to thirteen microns in diameter, and the term macrocyte to those of nine to eleven microns. Schistocytes are fragments of red cells. Gigantocytes are more than twelve microns, and may be as much as twenty microns in diameter, in the stained dried smear. Chlorotic cells are edematous cells. They are not pathognomonic of chlorisis but may occur in almost any form of anemia. Demilune bodies occur in red cells which have imbibed water. In such cells the stroma containing hemoglobin occupies a narrow area along one edge of the periphery of the swollen cell, the form suggesting that of the new moon. They are found in chlorosis and in severe anemias due to long malnutrition. Sickle cells are peculiar comma-like or sickle-shaped red cells found in the blood in certain anemias of the Negro race. Poikilocytes are red cells of abnormal form. Anisocytes ae red cells of abnormal size. The term as generally employed is anisocytosis, that is, a state of the blood characterized by great variation in size of the red cells. Nucleated red cells present similar variations with similar names. Microblasts are nucleated microcytes. Macroblasts or megaloblasts are nucleated macrocytes or megalocytes. Normoblasts are nucleated normocytes. Erythroblasts are nucleated cells containing hemoglobin, but rather larger than normoblasts; the term is commonly applied to cells intermediate between megaloblasts and normoblasts. Poikiloblasts ae nucleated poikilocytes. Many other terms have been employed by different authors, but these are explained by the author who so employes them and require no further discussion. To some extent these variations in the forms of red cells are due to their imperfect development; this condition is especially true in the primary anemias. Secondary anemias are characterized by cellular developmental imperfections after the red bone marrow has been seriously affected. In other conditions the red cells are only extremely fragile, being normal or about normal in size and form within the blood vessels, but becoming greatly distorted while the blood is being prepared for examination. In still other conditions the abnormal condition of the plasma causes changes in the form and size of the red cells, so that cells originally about normal become abnormal in size and form while still in the circulating blood. Abnormal chemical relations within the red cells may cause marked variations in their affinity for stains; this condition is called polychromasia. Poikilocytes and microcytes are especially abundant in the secondary anemias, chlorosis and sickle-cell anemia. Anisocytosis, polychromasia and megaloblastosis are most common in pernicious anemia and in certain forms of secondary anemia, notably those due to lead poisoning, certain intestinal worms and late stages of cancer with metastases in the bone marrow. Blood in which the average erythrocyte contains less than the normal amount of hemoglobin is said to be characterized by hypocytochromia, while blood in which the amount of hemoglobin in an average erythrocyte is above normal is said to be characterized by hypercytochromia. In hypocytochromia the color index is below one; as in most secondary anemias and notably in chlorosis. In hypercytohromia the color index is above one, notably in pernicious anemia and in certain anemias due to intestinal parasites. These red cells are always abnormally large. Red cells appear to be extremely fragile; they change shape readily and are rather easily laked in vitro. Yet in many respects they present remarkable stability in form and functions. Blood is often taken from one individual and placed
in the veins of another who needs blood. This transfusion, as it is called,
is part of the ordinary treatment in severe anemias and after severe hemorrhage.
The transfused blood cells retain their form and their functional power
for some days, and in one of our cases transfused cells were recognizable
for three weeks after having been place in the veins of the recipient.
That the transfused cells are able to carry on their proper functions in
the blood vessels of the recipient is shown by the immediate relief in
symptoms which occurs after the transfusion, and also by the fact that
the nucleated, reticulated and other immature erythrocytes often disappear
from the peripheral blood of the recipient very soon after the transfusion
of new blood into his veins.
In slightly thick smears of blood there occurs a peculiar grouping of the red cells in such a way as to suggest a pile of saucers or bowls, or a roll of coins. This tendency seems to be purely physical phenomenon, partly due to the peculiar shape of the erythrocytes, and partly to their stickiness and their lack of any true cell wall. Rouleaux once formed are not necessarily permanent. In slides examined on the warm stage rouleaux often form normally, then the cells break apart and rearrange themselves, sometimes in islet-like piles, sometimes in other rouleaux. Sometimes the rouleaux formation is complete within one minute in normal blood; sometimes it is delayed for three to fifteen minutes after the blood is taken. Sometimes rouleaux are not formed at all, or are very short and include only a small proportion of the cells present. Abnormal conditions often affect rouleaux formation,
and this may occasionally be helpful in diagnosis. In ordinary anemias
the rouleaux are subnormal; in pernicioius anemia they are almost or quite
normal. In pyogenic states they are usually normal; in cancer they are
either very much delayed, are subnormal, or, being formed, break apart
and the red cells form islets instead of rouleaux. Rouleaux are subnormal
or absent in Hodgkins disease and are usually subnormal in the leukemias.
The resistance of the red blood cells to hypertonic, hypotonic and hemolytic solutions has been studied with much care. In cases of doubtful diagnosis tests of the resistance of the blood cells may give pathognomonic facts. The manner in which the hemoglobin is held in solution is not definitely known, but various agents which cause laking are recognized. Saponin acts by injuring the lipoid substances of the stroma. The resistance of red cells to solutions of saponin remains within normal limits in pernicious anemia, diabetes and exophthalmic goiter, and is considerably increased in the secondary anemias, in syphilis, tuberculosis, splenomedullary leukemia, polycythemia vera and in normal or abnormal blood after splenectomy. The resistance of the red cells is considerably diminished in hemolytic jaundice, both to solutions of saponin and to hypotonic salt solutions. Normal red cells show no hemolysis until the salt in a solution is diminished to 0.44% or less, and complete laking rarely occurs, in normal blood, until the salt is diminished to about 0.34%. In hemolytic jaundice, on the other hand, the cells begin to lake in salt solutions of 0.6% or sometimes 0.7%, and complete laking occurs at 0.4% or even 0.45%. In blood which contains recognizable amounts of bile
pigments but not bile salts laking does not occur on the warm slide, but
blood which contains bile salts or acids as well as bile pigments shows
laking on the warm slide within five to fifteen minutes.
When the blood of one person is transfused into the veins of another the results upon the recipient may be good or bad according to whether the two bloods are compatible, that is, whether there is any agglutination of the cells of either blood by the plasma of the other or not. Hemolysis may or may not occur with agglutination, but it does not occur without agglutination. The human race is divided into four groups according to the relative compatibility of the blood of different groups, and it is most important that transfusion is not employed until the group relations of recipient and donor have been established. Very young infants are universal recipients, and it is usually safe to use transfusion for them without grouping. It is, indeed, impossible to determine to which group a very young baby belongs. The four groups depend upon the fact that the serum contains one or both of two agglutinins, and that the red blood cells contain one or both of two receptors. It is not possible to find an agglutinin in the same blood with its corresponding receptor, since such a relationship would not permit life at all. Group I, Jansky, or Group IV, Moss. This group includes about 40% of all Caucasians. The cells of these individuals are not agglutinated by the serum of other groups. The serum from this group agglutinates the red cells of all the other three groups. Group II, Jansky, or Group II, Moss. This group also includes about 40% of all Caucasians. The serum of this group agglutinates the cells of Group III, Moss and Jansky, and of Group IV, Jansky or Group I, Moss. The cells of this group are agglutinated by the serum of Group I, Jansky, or Group IV, Moss; and by the serum of Group III, Moss or Jansky. Group III, Jansky, or Group III, Moss. This group includes about 10% of all Caucasians. The serum of this group agglutinates the cells of Group II, Moss or Jansky, and of Group IV, Jansky, or Group I, Moss. The cells of this group are agglutinated by the serum of Group I, Jansky or Group IV, Moss, and by the serum of Group II, Moss or Jansky. Group IV, Jansky or Group I, Moss. This group includes about 10% of all Caucasians. The cells of this group are agglutinated by the serum of all three of the other groups. The serum of this group exerts no agglutinating influence upon the cells of other groups. The following hypothesis explains the grouping fairly adequately: Individuals are placed in groups according to the behavior of their cells with the serum of other groups. If the cells contain both types of receptor the serum from all other groups must agglutinate those cells. Conversely, the serum of this group cannot contain any agglutinin, since life would be impossible under such circumstances. This person belongs in Group I, Moss, or Group IV, Jansky. If an individual has serum which contains both types of agglutinin it is evident that his cells cannot contain either type of receptor, since such a condition would be incompatible with life. His serum agglutinates the cells in all three other groups, and he belongs in Group I, Jansky, or Group IV, Moss. If an individual has serum containing the A type of receptor and the beta type of agglutinin his serum agglutinates the cells containing the alpha type of agglutinin; he belongs in Group II, Moss or Jansky. If an individual has red cells with the B type of
receptor and the alpha agglutinin is found in the serum of his blood, then
he belongs in Group III, Jansky or Moss.
The behavior of the red cells with reference to agglutination is a constitutional trait which follows Mendels Law as a dominant characteristic. Babies under about ten weeks of age do not seem to possess agglutinins or receptors and are therefore included as universal recipients. After about three months of age the baby can be grouped and this grouping seems to be permanent throughout life, though there may be changes in the vigor with which the cells are agglutinated or with which the serum agglutinates other cells. Other than Caucasian races show different group relations. Any primitive race usually belongs to a certain group, but different primitive races may belong to different groups. Mammalian species may be classified upon a basis of blood-grouping and this method promises good results in biological studies. The blood of fourteen chimpanzees was tested with the blood of human groups; all belonged in Group II of the human groups. Many interesting relations have been reported for different mammals. The technique of blood transfusion is beyond the
scope of this discussion. The underlying principles are of interest in
any discussion of the red blood cells.
A recipient of one group may safely receive blood from a donor of another group provided the donors cells are not agglutinated by the serum of the recipient. The serum of the donor is so greatly diluted that agglutination of the recipients cells probably does not occur. Therefore the individuals of Group IV, Moss, or Group I, Jansy, are generally considered universal donors. The cells of this group are not agglutinated by the serum of any group. Since the serum of Group I, Moss, or Group IV, Jansky, does not agglutinate any cells, these individuals may receive blood from any group; they are considered universal recipients. Before transfusing blood both donor and recipient must be grouped. Even within the same group, occasionally, or when donor or the recipient are of a universal type, the two bloods should be cross-matched. This is because there are individuals which do not belong exactly into any one of the four groups, and also because of the possibility of error in the preliminary grouping. It is true that errors are rare, but their possibility must be considered. If second or later transfusions are necessary the
tests must be repeated even though the same donor be employed. This is
because it occasionally happens that after the first transfusion the recipient
may develop an anti-body for the blood of the donor, in which case serious
reaction may occur after the second transfusion. In any case the blood
should be given very slowly during the first ten minutes of the transfusion
and if any adverse symptoms occur the procedure immediately terminated.
Normal adult human blood contains from 4,500,000 to 5,500,000 erythrocytes per cubic millimeter. Rarely the number may reach 6,000,000 per cubic millimeter; as, for example, in a young man of good physique who has been engaged in violent exercise. Welkers original estimate, in 1854, of 4,500,000 for women and 5,000,000 for men is still accepted generally as correct, though these figures appear to be too low by about a half million in each case. The figures vary considerably under normal conditions, and to a very great extent in abnormal conditions. Normal may be considerable, and it is of great importance that blood counts should be made with proper control of these factors. The blood of women is generally supposed to be about 500,000 red cells poorer than the blood of men. This is not really a sex trait, however. It is due to the differences in the lives of the sexes from the beginning of puberty. Blood counts made of men and women of all ages, whose life habits are similar, show no differences which can in any way be considered due to sex. Students, teachers, clerks or bookkeepers whose lives are spent indoors and whose muscular activities are slight have about the same blood counts and hemoglobin percentages, whether they are men or women. On the other hand, counts made of women whose lives are spent in active, muscular work out of doors have blood counts which do not vary from those of their brothers. In Southern California, for example, women living alone or by twos or threes sometimes attend to the work of small ranches alone. Counts made of the blood of such women follow that usually found in men in similar occupations. Since women do, generally, live less active lives and are more closely confined within doors, a difference of about a half million erythrocytes to the cubic millimeter is expect ed. In estimating the percentage of the normals the occupation and not the sex are to be considered. Very few actual counts of early fetal blood have been reported. The number of red cells reaches its highest normal point very soon after birth, then diminishes until just before puberty. The count then rises gradually, modified by the other factors presently to be mentioned, until old age. The anemia due to senile disturbances then may cause a fall; but it is probable that old age in itself is associated with a diminution of the water, and thus with a continually rising count. The following table is compiled from counts reported from several laboratories, together with the records from the clinics and laboratories of The Pacific College of Osteopathy in Los Angeles, and The A. T. Still Research Institute in Chicago and Los Angeles. Children
The red cell count may vary in blood taken from different parts of the body. Local hyperemia causes local rise in the red cell count. The count is higher in dependent parts of the body. If one hand is elevated and the other hand allowed to dangle, the count is higher in the blood taken from the dependent hand and lower in the hand which is held higher than the body. If the hand is supported in an elevated position the count is higher than if it is held upward by muscular activity. Exercise of any part of the body increases the cell count in that region. If the arm and hand are vigorously exercised, the blood taken from a finger shows a higher count than blood taken before the exercise. If one arm only is exercised, the blood taken from a finger on that side has a higher count than blood taken from a finger on the opposite side. The constriction of a limb, as by an elastic band, causes increased red cell count. Any part of the body in which the circulation of the blood is delayed has a higher red cell count than normal areas. Very vigorous exercise involving a large part of
the entire musculature increases the red cell count, sometimes by 1,000,000
cells or even more.
Temperature changes have an effect only if somewhat prolonged, or if associated by marked changes in humidity. Generally speaking, the red cell count of nearly all normal individuals is about 500,000 higher in winter than in summer, in climates with marked seasonal changes. Persons who leave cold regions to go to the tropics may lose a million or more of red cells per cubic millimeter; persons who go from very warm to cold climates gain red cells in about equal numbers. No doubt this variation associated with changes in temperature is due, in part, to the same factors which cause variations due to altitude, that is, to changes in muscular activity, appetite, respiration, pulse rate, blood pressure and other functions which vary directly or indirectly as a result of diminished oxygen supply to the tissues. Hot baths, especially with much sweating, increase the red cell count. Cold baths decrease the red cell count at first, but after the reaction the count may be considerably increased. In typhoid fever cold baths may increase the blood count by almost or quite 2,000,000 cells. This increase disappears within an hour, or two hours at most. Local applications of heat and cold differ. Heat,
cold or rubefacients which increase the caliber of the peripheral vessels
increase the erythrocyte count in the affected areas; agents which lead
to vaso-constriction dimish the erythrocyte count. These variations are
due to the fact that the layer of plasma lining the capillaries remains
fairly constant under normal conditions; thus, when the caliber of the
vessels is increased, it is chiefly the cell-containing central area of
the capillary which is affected. Since the blood taken for counting comes
from the capillaries, the variations in the counting due to these conditions
may reach 200,000 or even more.
The person who travels fairly rapidly from lower to higher altitudes shows increased red cell counts, about 100,000 cells for each 2,000 feet in elevation. If he descends at once the count returns to its original number within one or two days. The change is so rapid that it seems impossible that increased hematopoiesis causes it, and there is no evidence of increased rapidity of blood formation in normal persons making the journey. Variations in the distribution of the blood, flushing of the rib marrow by rapid respiration, increased rapidity of evaporation of water from the skin and the lungs; fragmentation of the red cells with retention of almost normal structure; a shower of red cells from the bone marrow in answer to the lowered oxygen tension of the higher altitude and diminished efficiency of the hearts action are some of the factors which may be concerned in this temporary polycythemia due to increased altitude. The fragility of the cells is increased with the increasing cell count and the albumins and globulins of the blood are diminished at the same time. The person who remains at high altitude usually shows a blood count normal to him at the lower altitude or only slightly higher, within a few months. However, the Indians in Mexico who live on high mesas have habitual polycythemia, according to counts reported by F. Ocaranza. The air is very dry in that location. PLATE I
In our laboratories blood examinations made at Los
Angeles for people who have descended from higher altitudes show polycythemia
during the first few days to
A sudden rise to high altitudes gives the most marked variation; though the increase is certain no matter how gradually the rise is made. The factors which seem to affect this rise are many:-- Increased evaporation of the water due to the high altitude is one factor, though this is now considered of minor importance. Variations in the hearts action, due to the lowered oxygen tension, certainly modifies the peripheral count. These two factors are probably responsible for the increase noted immediately after the rise; the actual increase in the number of erythrocytes in the entire blood is due to other factors as well. The diminished oxygen tension of the higher altitudes is directly or indirectly responsible for the permanent and actual increase of the cells, and this acts upon the body in several ways: Increased need of the body for oxygen leads to increased hearts action, and thus to increased cell counts; Increased respiratory and cardiac activity means increased exercise; thus also increased cell counts; Increased respiratory activity means increased rib movement, this means increased efficiency of the circulation through the ribs (which include a great proportion of the red bone marrow of the entire body) with resulting increased nutrition of the blood-forming organs; Increased activity of the thorax with increased activity of the nerve centers associated with the respiratory and cardiac movements, causes increased activity of the nerve centers associated with the control of the hematopoietic marrow; Increased respiratory movements, together with the increased activity and appetite, due to changes of environment, may often be partly responsible for increased nutrition and thus for increased erythrocyte formation. This factor is not an important one in the variations noted under experimental conditions. It should be noted that in animals the variations in erythrocyte count are less marked and less constant than is the case with human subjects not under experimental conditions. Anemias due to hemorrhages, either experimental, accidental or pathologic, tend to more rapid recovery in high altitudes. The indirect effects of mountain climates,--the increased
activity, increased appetite and other effects of environmental change,--are
worthy of consideration in estimating the effects upon the blood of nervous,
over-tired or other patients who may be sent to elevated resorts for the
sake of erythrocyte stimulation. In this connection it should be noted
that the altitude does not always give increased powers of erythrocyte
formation. Patients who suffer from primary anemias or serious nutrition
or circulatory diseases may be harmed by changing to the higher altitude,
which makes upon their hematopoietic organs a demand greater than can be
met.
There is some tendency for the red cell count to drop during pregnancy, and this condition may be so exaggerated, in women who appear normal otherwise, as to simulate pernicious anemia. We have records of one woman in whose case each of three pregnancies was associated with decrease of the red cell count to less than 2,000,000, with color index at one or slightly above one. After the birth of each child the count rose to normal within a few weeks. The children were apparently perfectly normal. Normal menstruation and normal lactation do not seem to modify either the number or the character of the erythrocytes. Exercise, emotional states, excitement of various kinds, anything which raises the systemic blood pressure, increases the erythrocyte count through the filling of the peripheral vessels. Anger and fright do not increase the red cell count in animals or in human subjects after splenectomy, nor in animals after the solar plexus or the splanchnic nerves have been injured. Anger and fright cause increased contractions of the splenic capsule and this may account for the increase in normal subjects. Anything which abnormally increases the action of the kidneys or which increases the secretions of other glands increases the erythrocyte count by lessening the water content of the blood. Anything which increases the respiratory movements increases the erythrocyte count, both immediately and permanently. Probably the immediate rise is in part due to rising blood pressure and increasing heart action, and partly to the flushing of the rib marrow and thus an increase in the rate at which the new cells are thrown into the general circulation. The permanent increase in the erythrocytes associated with habitual increase in the respiratory movements is partly due to the better nutrition, and partly due to the better circulation through the red bone marrow of the ribs. Eating a light meal may not modify the erythrocyte count perceptibly. After a heavy meal, the increase in the water thrown out with the digestive secretions lowers the watery content of the blood, and thus raises the erythrocyte count. Later, the increased absorption of the products of digestion causes a lowering of the erythrocyte count. A later rise may occur, due to either an increased formation or to a flushing out of the cells already formed. This later increase is not usually seen in the blood of normal persons but it may be quite marked after a period of starvation or other malnutrition. The red cell count is somewhat higher at about two oclock in the afternoon in human subjects. This rise rarely exceeds 50,000 cells per cubic millimeter. It occurs without regard to fasting or to eating at noon. Persons who habitually eat nothing at noon, those who eat heartily at noon, those who habitually eat but who omit one noon meal, those who habitually omit the noon meal but who take food for one day only, all show about the same rise in red cell count within half an hour or so of two oclock in the afternoon. A less marked rise occurs at about the same hour in the morning. Transient polycythemia may be caused by any condition which concentrates the blood or which causes marked capillary or venous congestion. Diarrhea, sweating of severe degree, prolonged vomiting, cyanosis, sudden cardiac inefficiency as in decompensation and several less common conditions associated with loss of fluid or with acute dilatation of blood vessels cause temporary increase in the red cell count of capillary blood. Erythrocytosis is a polycythemia which is a reaction to some increased demand for red cells, beyond normal limits. The term is analogous to leucocytosis. High altitudes to which the reaction is inefficient may cause such an erythrocytosis. Heart disease is especially important; in congenital heart disease the count may be very high. In a case in the clinic of The Pacific College of Osteopathy a count of 8,000,000 cells per cubic millimeter was found in a boy eleven months old, with congenital heart disease. In adults cardiac disease sometimes causes very high counts. In one of our cases three days before death the count was 7,000,000 cells. Lung diseases usually increase the cell count, although if such diseases persist anemia supervenes later. The red cells are increased during fevers, probably on account of the concentration of the blood. The red cells may be increased during rapid leucocytosis, as if the increased activity of the leucotopoietic centers raised at the same time increased activity of the erythrocytopoietic centers in the bone marrow. This relation is not invariable. The red cells are generally increased in epidemic encephalitis and in many brain injuries. Rabbits which have been given brain lesions show increased red cell counts. In patients with rupture of the spleen, or with disease involving a considerable part of the spleen and in animals and in human subjects after splenectomy, the red cell counts may be very high, even to 10,000,000 per cubic millimeter or more. Certain drugs and biological preparations increase
the red cell count, but the repeated use of such drugs is always followed
by a very persistent anemia. Other drugs diminish the red cells immediately
and permanently. Radium and X-ray also diminish the blood cells.
The term erythremia, in a manner similar to the term leukemia, applies to a disease of the blood forming tissues characterized by very greatly increased red cell counts. This increase is not due to any demand on the part of the tissues of the body, so far as can be determined, but to a primary disease or developmental defect of the hematopoietic system. The red blood cells are normally being formed and being destroyed continually. They undergo fragmentation and probably hemolysis in the circulation. Phagocytosis of the fragments and of elderly entire red cells occurs continually in the endothelial cells of the liver, spleen and bone marrow. Regeneration occurs normally only in the red bonemarrow, in the adult. It is very evident that if regeneration exceeds destruction, though only by a narrow margin, polycythemia must inevitably occur. If destruction exceeds regeneration by even a narrow margin, anemia must occur as inevitable. Normally the balance is maintained by increased regeneration when any unusual destruction of red cells occurs, and by diminished regeneration and by increased phagocytosis when the number of red cells tends to become too great. Only under distinctly pathological conditions does the number of red blood cells vary beyond the limit of efficiency of their oxygen-carrying function. The rate of development of red blood cells in the
human adult has been variously estimated. Since the bile and urinary pigments
are derived from hemoglobin, the measure of these pigments in the urine
and the feces should indicate the number of red cells destroyed each day,
and thus the number of new cells formed.
Women are able to lose from fifty to five hundred grams of blood at each menstrual period, yet they show little or no increased erythrocytopoieses before the menstrual period and no erythrocytopenia afterward. This loss, then, must be relatively negligible when compared with the normal formation of new cells. After single hemorrhages amounting to five hundred grams of blood no abnormal conditions are recognizable in the peripheral blood. The removal of one hundred to three hundred grams of venous blood daily except Sundays for three weeks, for experimental purposes, caused no recognizable changes in the hemoglobin or the red or the white cell counts and no appearance of immature red or white cells, in one case in our records. After comparing all the reports accessible with our own records, it seems that many normal women must form about ten thousand cells each second all the time in order to provide menstrual blood alone. Various normal and abnormal conditions increase the
rate of blood formation, and thus the number of cells in the peripheral
blood.
In early embryonic life the first red blood cells appear in the extra embryonic mesoderm. There is a possibility that entodermal cells emigrate to this location. Groups of cells which are sometimes called blood islands appear, and in the midst of these there are certain cells which become flattened and somewhat elongated These form the endothelium of the blood vessels. The cells within these embryonic vessels develop into red corpuscles. The cells without the vessels ultimately form leucocytes and adventitial cells, and the muscular walls of the blood vessels. After the first cells are formed in the extra embryonic mesoderm, other tissues within the embryo begin to form blood cells, and during embryonic life both red cells and white cells seem to be formed almost anywhere. Later the liver and the spleen are sites of abundant hematopoieses and finally the red bone marrow assumes the most important place in the manufacture of red cells. After birth for a short time the spleen still provides some red cells, but the red bone marrow is pre-eminently the location of the hematopoietic tissues after birth. In normal adult human life the red cells are all
formed in the red bone marrow. The origin of the normoblast, in normal
adult human blood, presents several problems. In the red bone marrow are
found the earlier cells of the erythroblast series. These are cells which
resemble small lymphocytes in some degree, but which contain a small amount
of hemoglobin within their protoplasm. The nuclei are vesicular and the
chromatin is in small and rather scanty masses; there may be one or two
nucleoli. This cell, when stained after Sabins method, may show several
mitochondria, usually of somewhat elongated form. This cell divides into
two daughter cells and these into others; within one to a few divisions
the daughter cells show more abundant hemoglobin, smaller nuclei with denser
chromatin and this arranged in typical cartwheel form; nucleoli are usually
absent and mitochondria very scant or absent. This nuclear structure is
characteristic; the chromatin is arranged in rather dense masses which
are especially abundant around the periphery of the nucleus. This causes
the cartwheel appearance of the cell as seen in a thin smear of blood.
The megaloblast nucleus has rather dense masses of chromatin but they are
not quite typically of the cartwheel structure. The erythroblast nucleus
shows the cartwheel arrangement clearly, as does the normoblast nuelcus
and the naked nucleus found in the normal adult red bone marrow and in
the blood of pernicious and certain other very severe forms of anemia.
The erythroblasts and the normoblasts contain fairly abundant hemoglobin,
and this increases with the maturity of the cell until the typical erythrocyte
hemoglobin concentration is reached.
The manner in which the nucleus of the normoblast is lost has been the subject of much discussion. In the laboratories of The A. T. Still Research Institute studies have been made of the hemocytopoietic tissues of human embryos of ten days, three week, ten weeks, four months and later development; of embryos of rabbits, guinea pigs, cats, rats, moles and gophers; of the red bone marrow of human subjects dying of acute diseases and of aplastic, pernicious and several forms of secondary anemia, and of the red bone marrow of adult rabbits, dogs, cats, rats, guinea pigs, gophers, and horses. The following method of discarding the nucleus seems certainly to be characteristic of human erythrocytogenesis. In other mammals it has been found present also, though in these the relations have not been quite so thoroughly studied as in the human. (Plate I) The normoblast has its spherical nucleus and its deep rim of hemoglobin-containing protoplasm. This nucleus being apparently efficient with its deeply-staining chromatin and its typical structure, is to be extruded. The nucleus shrinks slightly and assumes an eccentric position in the cell; it approaches the periphery of the cell gradually; the protoplasm becomes thinner and thinner over the nucleus and finally shrinks away from it altogether, so that the nucleus is left altogether outside of the protoplasm, which is then an adult red cell, ready for its functional existence. Cytologically, of course, it is really in a senile state. The naked nucleus then swells slightly and soon it shows a thin rim of basophilic protoplasm around it. This protoplasm increases in thickness and develops hemoglobin within it. After a time the nucleus is again at the center of a normoblast and again the nucleus may be extruded in the same manner. It is not possible to say how many times a single nucleus may undergo this series of changes. In another series the nucleus of the normoblast has
a less definite structure. It stains less vigorously; the chromatin masses
are not quite typically cartwheel in arrangement; their outlines are less
sharply marked and the nucleus is a trifle larger. The nucleus swells somewhat,
its chromatin masses stain less and less vigorously; the nucleus loses
its sharply defined outlines; there is a clear area in the adjacent protoplasm;
the protoplasm accumulates hemoglobin most abundantly at the periphery
of the cell and finally the nucleus seems to dissolve within the red cell,
leaving some of the nuclear remnants called nucleoids or various other
names already mentioned in connection with the intracellular structures
of the erythrocytes. The extrusion of the vigorous nucleus and the digesetion
of the senile nucleus offer two parallel modes of conversion of the normoblast
into the erythrocyte.
During severe anemias the red cells do not follow the normal course of development. The megaloblast may pass directly into the circulation. It may lose its nucleus prematurely and the large cell thus formed has scanty hemoglobin and is a megalocyte. The normoblast nuclei may fail to grow before division and microblasts be allowed to reach the blood stream; or the microblast may lose its nucleus and the microcyte pass into the general circulation. Protoplasm may be budded off from red cells, under abnormal conditions, and microcytes thus be produced. The cells may be so poorly made that they undergo distortion with facility; many poikilocyt es and poikiloblasts are so formed; others are improperly formed in the marrow and pass into the blood stream unchanged. In pernicious and other very severe anemias and in the leukemias, the immature blood cells may be so abundant as to cause the blood smear to resemble a smear made from red bone marrow or from pus. Under very abnormal conditions in the adult, red blood cells may be formed in other tissues of the body, as in embryonic life. The spleen, the hemolymph glands, areas of endothelial cells and connective tissues associated with areolar and fatty tissues, even the secreting glands of the body, occasionally show structures resembling red bone marrow, in which hematopoiesis is seen to be very active. Metastases from the red bone marrow occur in certain leukemias and in pernicious anemia, and it is often difficult to determine whether any given area of hematopoietic tissue found in some aberrant location is a metastasis or is an instance of metaplasia. Red cells which have passed their period of usefulness
must be quickly removed from the circulation, because it is only very rarely
that such cells are found in normal blood. There is a form of basophilic
degeneration which occurs in cells which are presumably too old to function
properly. In blood kept in vitro the red cells become fragmented, laked
or swollen according to environmental conditions but they do not become
basophilic. In the circulating blood in cases of jaundice and of intestinal
toxemia fragmentation occurs more abundantly than in normal blood. Fragments
and abnormal cells of various types are taken up by the phagocytic endothelial
cells (Kupffer cells of the liver; endothelial cells of the spleen, lymph
nodes and bone marrow) and by these cells they are digested. The processes
are not yet well understood, but the hemoglobin is certainly broken into
an iron-free pigment (bilirubin, hematoidin or some related compound) and
this pigment is eliminated in the bile and the urine. There is good reason
for believing that the reticular cells perform the major part of the work,
and that the true hepatic cells pass the pigments onward into the bile,
perhaps after elaborating them in some manner. Much further work must be
done before the steps of this procedure can be described, and the problems
presented by the dissolution of the red cells and the conservation of the
iron-containing molecules are very complicated.
The amount of hemoglobin carried by each red blood cell, the amount carried by a given amount of erythrocyte stroma, and the functional efficiency of the hemoglobin are subjects of some interest. These factors are determined by a study of the color
index, the volume index and the saturation index of the red blood cells.
This is the fraction obtained by dividing the hemoglobin percentage by the erythrocyte percentage. The hemoglobin and the erythrocyte count should be made, and the percentage of the normal for the age of the patient computed. The color index expresses the relative amount of hemoglobin carried by an average red cell of the patients blood. For example, a young baby should have 150 to 200 grams of hemoglobin per liter, according to its age; it should have 5,500,000 to 6,500,000 erythroytes per cubic millimeter. If these figures were given by a babys blood its color index would be 1., which is normal. An adult should have about 140. grams of hemoglobin per liter, and 5,000,000 erythrocytes per cubic millimeter. These figures give a color index of 1. An adult who has suffered a severe hemorrhage from accident, may, immediately after the loss of blood, give 60% hemoglobin and 60% erythrocyte count; 60% divided by 60% =1. which indicates that each erythrocyte is carrying the normal amount of hemoglobin, though it is evident that the number of the erythrocytes is diminished. The next few weeks usually alter the erythrocytes, if the hemorrhages are repeated. The same person may then have 55% hemoglobin and erythrocyte count of 110%. 55% divided by 110%=0.5 which is the color index characteristic of secondary anemia. The erythrocytes are then normal, or above normal, in number, but they carry about half the normal amount of hemoglobin. The blood-forming organs are reacting to loss of blood by manufacturing increased numbers of erythrocytes, but the hemoglobin is insufficient to form perfect erythrocytes; hence the low color index. In chlorosis extremely low color index is the rule. One case of ours gave an erythrocyte count of 120%; hemoglobin percentage of 30; 30% divided by 120%=0.25, the color index. In pernicious anemia, the color index is high, though the hemoglobin is very low. One case gave hemoglobin, 20%, erythrocyte count 15%. 20% divided by 15%=1.33, the color index. The color index may reach 3.2 in pernicious anemia. Nearly all secondary anemias show a low color index, usually below 0.7 and sometimes below 0.4. Anemias due to intestinal worms, especially bothriocephalus latus, often show high color index. Lead poisoning and most cases of pernicious anemia are characterized by high color index. Rarely in these cases the color index may be below unity. A patient with pernicious anemia or with lead poisoning
may suffer from some cause of secondary anemia, with resultant leucocytosis
and increased red cell count with low color index. The blood picture may
be very puzzling in such cases. After the acute condition has passed the
pernicious anemia type of blood picture recurs.
The red cells make up approximately one half the total volume of the blood. The volume index is the fraction secured by dividing the volume per cent of red cells by the red cell count expressed in per cent of normal. For example, if a patient has 5,000,000 red cells per cubic millimeter, or 100%, and if the hematocrit estimation of the blood cell volume is one-half the total blood volume, or 100% of the normal, the volume index is 100% divided by 100%, or 1, which is normal. If the patient has 4,000,000 red cells per cubic millimeter, or 80% of the normal, and has a cell volume of four-tenths of the total blood volume, or 80% of the normal volume, then also his volume index is 1.(80% divided by 80%). If the red cell count if 4,000,000 and the volume of cells is three-tenths the total blood volume, or 60% of normal, then the volume index is 60% divided by 80%, or .75. In other words, each red cell is about three-fourths of the normal volume or size, and this is the condition in ordinary secondary anemia. If the red cell count is 2,000,000 or 40% of the normal, and the cells make up one-fourth the total volume of the blood, or 50% of the normal, then the volume index is 50% divided by 40%, or 1.25. This is the condition characteristic of pernicious anemia, in which the red cells have an average volume of considerably more than normal. The volume index has about the same clinical significance
as the color index.
The color of the blood is due to hemoglobin. This remarkable substance is greenish yellow by transmitted light, and red by reflected light. The particular shade of red depends upon the chemical relations of the hemoglobin. The hemoglobin of venous blood, reduced hemoglobin, has a purplish tint within the veins or in a glass vessel into which it has been drawn without being exposed to the air. Oxyhemoglobin has the hue of arterial blood. Venous blood immediately takes up oxygen when it is exposed to the air, thus becoming oxyhemoglobin and assuming the characteristic brilliant scarlet tint. On exposure to carbon monoxide the hemoglobin unites with this gas, forming carbon monoxide hemoglobin and assuming a bright cherry-red color. The hemoglobin is carried in the erythrocytes. The
amount carried by each red cell varies somewhat in health, and varies within
wide limits under abnormal conditions. For example, in chlorosis the color
index may be as low as 0.2, which means that the average red cell is carrying
about one-fifth as much hemoglobin as normal red cells carry. In pernicious
anemia, on the other hand, the extremely large red cells may carry more
than three times the normal amount of hemoglobin.
The manner in which the hemoglobin is held within the meshes of the stroma is not definitely known. The stroma at the periphery of the cell is in a condition of slight tension, barely perceptible by delicate methods of examination. This peripheral area has been called a cell-wall though this is really a misnomer. Fragmentation of the erythrocytes does not cause laking, hence the peripheral tension is not the essential factor in keeping the hemoglobin within the stroma. The meshes of the stroma are rather coarse and the framework is open, so there is no actual mechanical restraint placed upon the hemoglobin. The hemoglobin within the erythrocyte is combined with sodium or potassium (sodium hemoglobinate; potassium hemoglobinate) and in this form it is much more soluble than in other forms. There is much reason for supposing that a very labile form of chemical union exists between the stroma and the hemoglobin, though just what the nature of this combination may be is not yet known. The study of hemoglobin is best made after the hemoglobin has been freed from the stroma. This process is called laking. The combination of hemoglobin and stroma is very stable under all conditions normal to the body, and even under many conditions which are distinctly abnormal. Certain abnormal conditions, not always in themselves very serious, may cause the hemoglobin to be set free within the blood vessels, thence to be eliminated by the kidneys, chiefly, and to some extent by the liver. Infections of several types may be directly or indirectly hemolytic. Infectious foci due to any type of the hemolytic streptococci may be apparently small and of negligible pathogenicity, yet the products of the activity within these foci may cause severe anemia. The malarial parasite causes fragmentation of the red cells which have been invaded and under certain circumstancese not yet well understood considerable laking of the blood occurs. The hemoglobinemia causes hemoglobinuria, the black-water of older writers. Syphilis may cause hemoglobinemia and hemoglobinuria, indirectly. Jaundice causes hemoglobinuria which may be severe but is often unrecognized on account of the associated choluria. The bile salts lower the surface tension, dissolve the cholesterin, destroy the structural relations of the stroma and set the hemoglobin free in the plasma. Bile pigments have less marked effects but seem to be somewhat hemolytic. Diseases of metabolism occasionally are associated
with hemolysis. Scurvy causes abundant hemorrhages, which in turn cause
anemia and hemoglobinuria. Intravascular laking occurs in severe cases
of scurvy. Severe frostbites are often followewd by hemoglobinemia and
hemoglobinuria. Extensive burns and insolation are also often followed
by intravascular laking. The local destruction of the blood cells is not
enough to account for the degree of laking, and there is good reason to
suppose that the poisonous products of the injured tissues destroy the
red cells within the blood vessels.
Paroxysmal hemoglobinuria is a peculiar condition in which hemoglobin is excreted occasionally in the urine, very often without any recognizable cause. Persons so affected do not seem seriously injured by the excretion of this important constituent of the blood. They may be somewhat weak after an attack, and occasionally an unusually severe attack may be followed by transient anemia. Attacks may occur after exposure to moderate variations in heat or cold; they are often precipitated by chilling of the surface of the body. Pre-existent hemoglobinemia is rarely demonstrable. The red cells of persons so affected are somewhat less resistant to the action of various hemolytic agents in vitro than are normal erythrocytes. Many poisons are hemolytic. Generally speaking, those poisons which are lipolytic are also hemolytic, and they act upon the blood cells by dissolving or injuring the meshes which make up the stroma. There are other poisons which are distinctly hemolytic but are not known to be lypolytic. Many drugs used in the treatment of disease are hemolytic, and the habitual use of such drugs is a cause of much chronic anemia. Tincture of iodine, potassium chlorate, arsenic, carbolic acid, napthol, ricin (in castor oil), benzol, lead compounds, sulphuric acid, hydrochloric acid and many of the coal-tar derivatives are a few of the poisons sometimes given as drugs which cause some degree of laking within the blood vessels. The venom of certain snakes and the poisonous substances of certain morels (toadstools) are also hemolytic. Transfusions of unfit blood may cause hemolysis.
This accident does not occur since the practice of studying the blood of
the donor in connection with the blood of the recipient has become common.
Laking of the blood is easily caused in vitro. The addition of distilled water, ether, any one of several salts in varying proportions, and many other substances cause the hemoglobin to be set free from the stroma. The stromata may be separated from the solution of hemoglobin by centrifugation and the hemoglobin removed for study. The stromata may be washed by adding water or other solvents to the sediment and centrifuging the mixture; the process can be repeated until the hemoglobin and such extractives as may be desired have been carried away by the supernatant fluid. The solution of hemoglobin has been studied in many laboratories. Hemoglobin is a conjugated protein with a formula which varies somewhat according to different chemists, but is about C758 H1203 N193 Fe S6 O218 with a molecular weight of about 16,669. Recent analysis seems to indicate that there are four atoms of iron instead of one atom in each hemoglobin molecule. Hemoglobin can be crystallized from the solution and the crystals so obtained vary for different animals. Related animals show some similarity of hemoglobin crystals, and this fact can be used as a basis for the classification of animal species. There are so many variations in the crystals due to variations in technique that medico-legal questions are not properly decided upon such evidence. Various analyses have been made of the hemoglobin
of different mammals. The exact figures vary, as do the figures given for
human hemoglobin, according to the methods of analysis which are employed.
The molecular weight of hemoglobin in the blood of many animals reported
varies from 14,780 to 18,370, with one atom of iron in each molecule in
each case.
Hemoglobin is easily broken down into two molecules, hematin and globin. Hematin makes up about four per cent and it contains all of the iron of the hemoglobin molecule. The globin makes up about ninety-six per cent and it contains all of the sulphur of the hemoglobin. Both these substances can be broken down into simpler molecules and these are often of physiological and pathological interest. Hematin is found in the spleen and the blood of patients with malaria. Hematin, injected into the veins of an animal, lowers the blood pressure, causes chills followed by fever, and causes petechial hemorrhages in the kidneys. Hemin crystals are formed by treating blood with glacial acetic acid; the procedure varies according to the amount of the crystals desired and the use which is to be made of them. Hematin can be made from the hemin crystals by treating them with alkalies. Hemin has the probable formula of C33 H3 O4 N4 C1 Fe. Hematin has a probable formula of C32 H30 N4 O3 Fe. Hematin can be treated with sulphuric acid and thus the iron is removed from the molecule. This iron-free pigment is known as hematoporphyrin, which has a deep purple color in solution. Its presence in the decomposed blood around bruises or other hemorrhagic tissues produces characteristic purple tinte. Hematoporphyria has the formula of C34 H38 N4 O6, and is isomeric with bilirubin. Both hematoporphyrin and bilirubin are present in the blood plasma and in the urine in traces, normally, and either or both may be increased under certain pathological conditions. Hematoporphyrin combines to form several closely related compounds. These are found in the urine of patients with rheumatism, several disorders of the liver, tuberculosis, malaria, and, occasionally, in other high fevers. The use of certain drugs, such as trional, sulphonal, veronal, the salicylates and many of the coal-tar products is sometimes followed by hematoporphyrinuria. Hematoporphyrin has a peculiar property of increasing the sensitivity of the skin to light. Patients with hematoporphyrinemia may be seriously injured by exposure to ordinary daylight and may suffer serious skin lesions as a result of exposure to sunshine. Hemofuscin is an iron-free brownish pigment derived from hemoglobin. The name has been applied to several quite different compounds by different investigators and hence has fallen into disuse. Hematoidin is an iron-free pigment derived from hemoglobin. It is often present in old accumulations of blood within the body. Its presence indicates that hemoglobin can be broken down by the cells and fluids of the tissues in various regions of the body, the iron-containing moiety being carried away to be used again and the hematoidin left with the clot, useless and not seriously injurious. Hematoidin is probably identical with bilirubin. Methemoglobin is isomeric with hemoglobin. It is
produced spontaneously in blood which has been allowed to stand for some
weeks, or it can be produced in vitro by the action of potassium ferricyanide
or potassium permanganate. Methemoglobin has a brownish color and it is
occasionally found in old hematomats.
Bronzed diabetes is a peculiar disease characterized by the deposit of hemosiderin and an iron-free pigment which is probably hemofuscin in the tissues of the body. The skin assumes a peculiar bronze-like tint not found in other diseases. Sugar metabolism is greatly disturbed and typical symptoms of diabetes mellitus may occur. Sulphemoglobin may be formed within the erythrocytes
by allowing hydrogen disulphide gas to pass through the blood whose coagulation
has been prevented. The anemia which is associated with intestinal putrefaction
is said to be due to the injury of the red cells by the hydrogen disulphide
produced by the putrefaction of the protein of the food in the small intestines.
Myohematin is the pigment of muscle cells. It is not quite identical in structure with hemoglobin but it performs the same function. It provides the muscle cells with nascent oxygen and thus facilitates the rapid oxidation processes essential to muscular activity. Myohematin forms a stable compound with carbon monoxide, as does hemoglobin, and is thereby rendered useless. The weakness and the marked fatigue on relatively negligible exertion which characterize chronic carbon monoxide poisoning are in part due to this fixation of myohematin. The digestion and absorption of the molecules thus destroyed is associated with considerable toxemia. The myohematin of the meat in food is a valuable source of the globin necessary for the formation of the hemoglobin of new blood cells. It must be remembered that the globin of senile blood cells is not saved, as the iron-containing part of the molecule is, and that fresh supplies of globulins are essential to the formation of the new supplies of erythrocytes constantly required by the body. The copper, magnesium, potassium and other inorganic elements needed for the synthesis of hemoglobin are also present in lean meat used as food. Several oxygen-carrying pigments are present in the body fluids of invertebrates. Hemocyanin or oxyhemocyanin occurs as a group of related compounds present in the blood or other body fluids of molluscs, cephalopods and crustaceans of fresh water and of salt water Compounds of the hemocyanin group are biological precursors of hemoglobin, though there is not any reason for supposing that hemoglobin was derived from any one of the group. In all the hemocyanin group copper has the place occupied by the iron in the hemoglobin molecule. The different compounds of this copper-containing oxygen-carrying pigment differ chiefly in the effects produced in their solutions by various reagents. For example a solution of hemocyanin derived from the snail becomes yellow on standing or after aeration with nitric oxide. A solution of hemocyanin derived from the lobster does not form the yellow compound even after long aeration with the same gas. Other animals produce hemocyanin which gives other reactions, hence there must be at least several different but closely related compounds included as hemocyanin. Copper does not seem as efficient as iron in the transportation of oxygen. Chlorocruorin is a green pigment found in worms and
a few marine animals. This pigment contains iron and it performs the respiratory
functions characteristic of hemoglobin in higher animals.
Both animals and vegetables contain many pigments which are concerned in the oxygenation and the oxidation processes of living cells, and in producing colors which have various functions. Chlorophyll and chromophyll are colored substances
occurring widely in the vegetable kingdom and in the bodies of certain
animals of very simple structure. Chlorophyll is especially efficient in
the utilization of the energy of the suns rays in the manufacture of certain
glucosides which in turn form starches and sugars in the sap of living
plants. These substances, in their pure form, do not contain iron, but
iron-containing molecules are always intimately associated with them and
it is by virtue of the iron-containing molecules that the cholorophyll
is able to build up the glucosides from carbon dioxide and water under
the influence of sunshine. The iron of these vegetable compounds is an
important factor in the synthesis of the hemoglobin of animals. Sulphur
and magnesium are other very important atoms associated with chlorophyll
and hemoglobin.
The total amount of iron in the whole body is only about six grams, and of this only about four-fifths are present in the hemoglobin. This iron is used very economically, over and over, in the formation of new blood cells. Very minute amounts of iron in the daily diet are sufficient. Restricted diets often fail to provide the globulins
necessary for the synthesis of hemoglobin but it is rare indeed that a
diet is too restricted to provide enough iron.
Diets rich in the globulins and albumins but relatively poor in iron are also efficient in supplying the elements required for regeneration. A diet of milk or of casein has little iron but animals are able to regenerate hemoglobin rapidly on such a diet, and patients with secondary anemia often recover speedily on a diet exclusively of milk. This fact is due to the presence of a fairly abundant supply of iron in the patients own tissues, which he is unable to utilize because of a lack of globulins and albumins. The milk diet provides these necessities in a form which is easily digested and absorbed and which avoids toxemia in considerable degree. Milk is not good as an exclusive diet for more than a few weeks because it does not contain all the substances necessary for the formation of hemoglobin nor for the maintenance of the needs of the body. Even in infancy an exclusive milk diet does not best provide for the needs of the growing body for more than a few months, at most, and babies seem to do better if other foods are given them at an early age. The anemia which occurs in children kept too long on an exclusive milk diet disappears rapidly when fruits and vegetables are added to the diet even though these may not contain any great amount of iron. The importance of adequate amounts of copper, sulphur, magnesium and potassium for the building of red cells should always be kept in mind. At birth the Kupffer cells of the liver are abundantly supplied with iron derived from the maternal blood. The blood, bone marrow and spleen of newly born babies all have a high content of hemoglobin and of other iron-containing compounds. This supply of iron is enough to maintain a normal amount of hemoglobin and myohematin for several weeks or months; that is, during the period normally associated with an exclusive milk diet. Except as animals eat other animals, the iron must
be derived from vegetables. Chlorophyll and chromophyll are iron-free vegetable
pigments which are closely associated with iron-containing molecules in
vegetable cells. Without the iron the chlorophyll is unable to utilize
the rays of the sun, carbon dioxide and water in the synthesis of the glucosides.
Chlorophyll is formed around the pyrrole ring which also is an important
part of the hemoglobin molecule. Vegetable cells containing iron, sulphur,
copper, magnesium and chlorophyll are an important source of the hemoglobin
and myohematin of the animal body.
Vegetables derive their iron from the earth in the form of ferric or ferrous salts. These are sufficiently soluble to be carried in the sap and to be utilized by the vegetable cells, especially the leaves, in the synthesis of those substances necessary to the activities of the chlorophyll granules. The oxygen-carrying function of the iron in these compounds is as important in vegetable economy as is the oxygen-carrying function of the iron of the hemoglobin of the animal. In the case of the vegetables the iron is necessary to the utilization of the energy of the sunshine in the synthesis of the glucosides. Iron does not seem necessary, however, in the further synthesis of the sugars and the starches from the glucosides. Even in the earth, before the iron is taken up by
the vegetables, its oxygen-carrying activities are economically useful.
There is a constant series of changing chemical relations of iron in the
earth. Varying combinations of iron with oxygen are especially noteworthy.
For example, ferrous silicate may be decomposed by carbonic acid into silica
and ferrous carbonate; this absorbs oxygen, liberates carbon dioxide and
forms ferric oxide. If decomposed organic matters are at hand, these take
up oxygen from the ferric oxide and ferrous carbonate is formed, to be
again oxygenated and decomposed. Sulphur and iron both act as oxygen carriers
in other groups of chemical reactions by means of which the oxidation of
organic matter is facilitated. Copper undergoes a similar series of oxidation
and reduction processes. A series of these alternating oxidation and reduction
processes, with nascent oxygen, is continually being set free in the earth.
Nascent oxygen is especially active in promoting oxidation. The growth
of bacterial food facilitates and hastens the decay of organic animal and
vegetable matter.
Variations in the amount of hemoglobin will be found in the number of red cells, which should be watched rather closely. Variations said to be due to sex are really variations due to habits of life. Men and women who are healthy and accustomed to active exercise or to hard work in the fields have blood with high hemoglobin content. Men and women who lead sedentary, indoor lives have blood with a low hemoglobin content. Since women usually are less active and usually spend more time indoors, the hemoglobin content of the blood of the average woman is about ten per cent less than the blood of the average man. Variations in hemoglobin due to age follow variations in the red cell count to some extent. The concentration of the blood immediately after birth is associated with very high hemoglobin content; from 180 grams per liter to 213 grams per liter, as reported from various laboratories. The red cell count is also very high at this time. The amount of hemoglobin diminishes gradually until about the fifth year, when the child may show only 110 to 115 grams per liter of blood. This is associated with a red cell count of about 5,500,000 per cubic millimeter. The red cells are smaller during childhood than they are in babyhood or during adult life. In any study of the blood of a child it is necessary to keep this peculiarity in mind, and to base the percentages of the color index upon the figures normal for the age of the patient. The following table gives the number of grams of hemoglobin per liter of blood found in Los Angeles in normal and practically normal individuals of different ages. The sexes are about equal. The determinations were made for fifty or more individuals in each group. Grams hemoglobin
Individuals between sixty and eighty years were considered
normal if no evidence of organic disease was noted. Naturally, these persons
were subject to some infirmities due to old age.
Daily variations follow variations in the red blood count. In order to secure logical statistics for comparison of the hemoglobin of different individuals or of the hemoglobin of the same individual under different circumstances it is necessary to examine the blood at the same time of the day. When the blood is examined at different hours the error due to this fact may be as much as ten per cent of the reported amounts. Variations in elevation cause variations in hemoglobin similar to the variations in the erythrocyte count due to the same conditions. On ascending to high altitudes the hemoglobin increases less rapidly than do the red cells. On descending the hemoglobin diminishes as rapidly as do the red cells. The color index thus is low during the few days following elevation and this is probably due to the rapid formation of new red cells. The color index remains unchanged during the few days after descent because new red cells are not formed rapidly. The increase in hemoglobin occurs in animals subjected
to diminishing oxygen tension, and in human subjects for whom the same
tests are made. For each fall of 100 millimeters of mercury in the atmospheric
pressure the hemoglobin increases about ten per cent.
In order that hemoglobin may be properly synthesized several conditions are necessary. The last steps in the building of hemoglobin take place within the erythrocyte stroma, and this stroma is essential to the process. It is not essential that the red blood cell be itself of normal size or form but it seems to be necessary for the meshes of the stroma to be at least approximately well built. The cholesterin, lecithin and other lipoids of the stroma are derived chiefly from the meat and other animal proteins of the food. Vegetable proteins are utilized by most people with some difficulty. Hemoglobin differs from chlorophyll in its relations to sunlight. Without sunlight chlorophyll is not developed. Hemoglobin can be developed in the entire absence of sunlight. It is true that moles, gophers, ground squirrels and other burrowing animals are exposed to the sunlight at least for short periods of time. Horses, mules and burros which are born in mines and live all their lives in darkness have normal hemoglobin averages. There are mines in which horses have lived for several generations in darkness which is complete except for the candles of the miners, and yet they have normal hemoglobin. The food must include some of the iron-containing molecules, and must contain an abundance of the protein suitable for providing the globin. It should be remembered that the iron-containing part of the hemoglobin forms only about four per cent of the hemoglobin molecule, and that the iron is preserved to be used over and over again; the globin, on the other hand, forms about ninety-six per cent of the hemoglobin molecule, and this is used as food for the body and as a source of various secretions; it is not, apparently, utilized even a second time in the building of hemoglobin. The necessity of an abundant supply of the proteins from which the globin part of the hemoglobin molecule can be synthesized is evident. The protein foods must be properly digested In ordinary conditions this presents little difficulty. In pernicious anemia the lack of hydrochloric acid in the gastric juice prevents adequate digestion of the proteins, and this seems to be an important factor in the pathogenesis of the disease. The severity of disease of the digestive tract does
not bear any direct relation to the degree of anemia produced by the disease.
Very severe ulcers of the stomach or the duodenum may be present in patients
whose hemoglobin is almost or quite normal; in other cases either a small
or a severe ulcer may be associated with marked anemia. Occasionally very
severe anemias, presenting marked resemblance to pernicious anemia, may
be associated with ulcers of the stomach which seem to be negligible in
extent. This relation does not always depend upon the presence or the absence
of normal hydrochloric acid secretion. Since patients with pernicious anemia
lack hydrochloric acid in the gastric juice, it is not always possible
to determine whether the gastric disease is the cause of an atypical anemia
resembling the pernicious type, or whether the condition is really Addisonian
anemia with the related achlorhydria. There is no reason for difficulty
in diagnosis when the blood picture is definitely megaloblastic or is definitely
of the secondary type.
Lesions affecting the digestive tract often cause a severe anemia of the secondary type. Lesions of the fifth thoracic vertebra, neighboring vertebrae and ribs are usually associated with hyperchlorhydria and ultimately with gastric ulcers. Anemia is not usually severe in these cases. Mild bleedings are always present in the stomachs of laboratory animals with the lesions mentioned but they do not usually become anemic. Probably the extravasated blood is digested and absorbed in these cases. Lesions of the seventh thoracic vertebra and neighboring vertebrae and ribs are associated with hypochlorhydria, gastric atony and malnutrition. Anemia is usually present in these cases but is rarely very severe. In animals it is always of the secondary type, but in human patients in whom the lesions have been present a long time there may be factors in the blood picture which suggest pernicioius anemia. Lesions of the tenth thoracic vertebra and neighboring vertebrae and ribs affect the circulation through the liver. The bile pigments and the bile salts are mildly toxic in small amounts. The bile salts cause laking of the blood in small degree even while the blood is circulating through the vessels. The hemoglobin thus set free is eliminated by the kidneys. The anemia due to this elimination of hemoglobin is not usually severe and is of the secondary type. In none of our cases has there been any evidence of a megaloblastic type of anemia associated with cholemia. Rib lesions are associated with disturbed nutrition of the local areas of red bone marrow. Without good circulation of the blood through the hematopoietic tissues, the hemoglobin is not normally provided for the red blood cells. When any considerable area of red bone marrow is affected by lesions the blood shows low hemoglobin, very low color index and many immature forms of red cells and of granular white cells. After correction of the lesions the red blood cells soon contain normal amounts of hemoglobin and the immature forms disappear. The relation of certain of the internal secretions
to the synthesis of hemoglobin has not yet been explained. It is, however,
true that patients suffering from certain forms of goiter and from diseases
affecting the ovaries or the testes often suffer from severe grades of
secondary anemia, and that with recovery of normal conditions of these
glands the hemoglobin returns to normal. The circulatory disturbances of
these glands, due to bony lesions, are often associated with anemia which
seems very much more profound than is warranted by the relatively mild
pathological changes in the tissues affected. On correction of the lesions
the hemoglobin returns to normal.
As the red cells become senile and inefficient they are removed from the circulation. They may become fragmented as they are being carried in the blood stream, and they are easily though not frequently recognized in smears made from the capillary or the venous blood. When there is increased destruction of the red cells, for any reason, the fragmented forms may be quite abundant in the capillary blood. The fragments of erythrocytes are ingested by the endothelial cells of the liver, spleen and other tissues, the hemoglobin is broken down into an iron-containing moiety, which is carried by the blood to the red bone marrow, and an iron-free pigment, which is transformed into bilirubin and related pigments. The Kupffer cells in the liver are especially efficient in the secretion of the bile pigments but the cells of the reticulo-endothelium system anywhere perform this function normally. When the liver of the human subject is diseased, or when the liver of a laboratory animal has been removed, the cells of the other parts of the reticulo-endothelial system may handle the hemoglobin metabolism with apparently perfect efficiency. The pigments of the urine and the bile are chiefly, if not altogether, derived from the iron-free pigment of the hemoglobin. These pigments are easily replaced from the food materials ingested. Hemoglobin set free in the blood plasma by the disintegration of the red cells is excreted from the body as such. The hemoglobinuria sometimes associated with severe forms of malaria is due to the destruction of the red cells by the malarial parasite with resultant hemogloginemia. Laking of the blood in the circulation as a result of poisoning by certain reptiles or by certain drugs is also associated with hemoglobinuria. Hemoglogin freed from the cells in stagnant blood, as in old hemorrhagic areas or in hematomata of long standing, undergoes various changes into methemoglobin, hematoporphyrin and other pigments. Ultimately these are absorbed by the blood and the lymph if there is any circulation of these fluids through the bloody accumulations, and they are excreted in the urine and the bile. Cells of the reticulo-endothelial system ingest the cells of extravasated blood in their immediate neighborhood, change the hemoglobin into an iron-containing and an iron-free moiety, transform the iron-free moiety into bilirubin or some related compound and allow the iron-containing molecules to be carried away by the plasma and the lymph, ready to be utilized again in the production of hemoglobin in new red cells in the bone marrow. This economical utilization of the iron containing part of the hemoglobin is not perfect, and there is a constant, though minute, excretion of iron in the bile and the urine. The iron found in feces is chiefly a part of the food which has not been absorbed into the body at all; the traces of iron found in the bile are excreted with the feces from the body. |
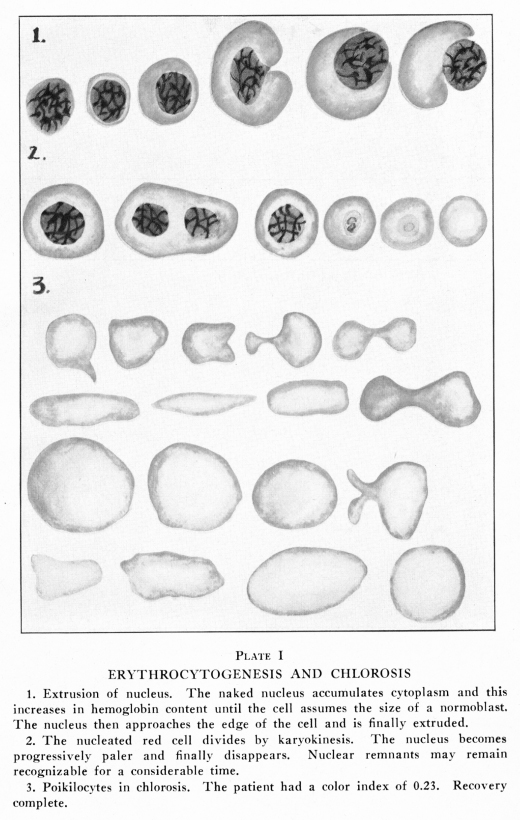{kind=link}
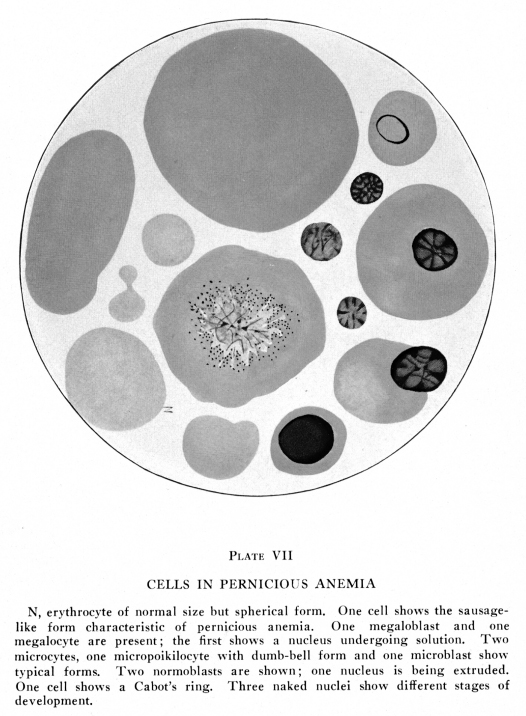{kind=link}
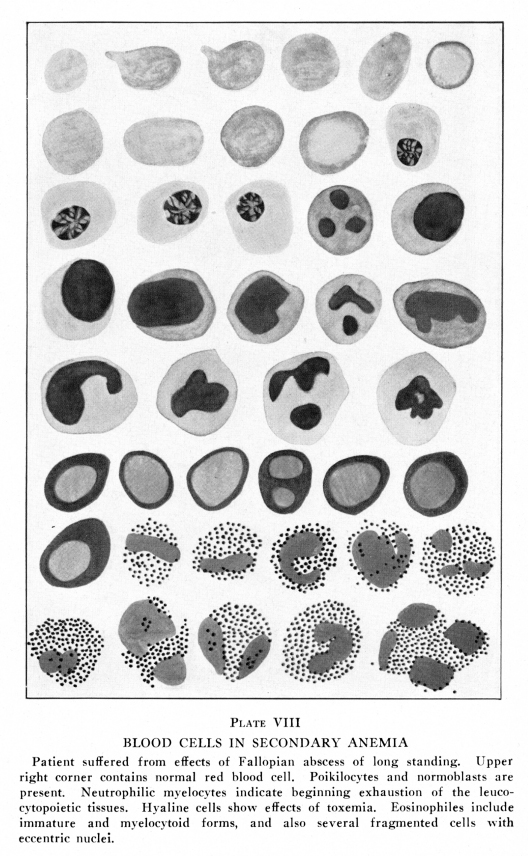{kind=link}