
|
The phylogenetic development of the hemispheres presents a series of puzzles. In man there seems no reason to doubt that the functional activity of the cortical neurons is essential to the occurrence of consciousness, as the term is usually understood. If any consciousness is possible otherwise, it must be of a character totally different from the phenomenon now recognized physiologically as consciousness. This possibility may be omitted from a physiological discussion. Phylogeny of Cerebral Functions Among animals in the vertebrate series the development of the cerebral hemispheres goes hand in hand with the development of reactions more and more adapted to the needs of the particular individual in the midst of a particular environment, rather than to the needs of one of a class in answer to the demands of an unvarying environment. The development of the cerebral hemispheres provides the structural possibilities for a great number of potential reactions in answer to any environmental demand. Thus, the possibility of individuality in the various activities of the individuals of any group of animals becomes fixed. Another result of the development of the cerebral hemispheres is found in increasing speed of reply to environmental variations. Variations in environmental conditions affect the life history of brainless animals chiefly or only through varying them, as a race, in perpetuating those individuals capable of making adequate reactions to those changes. But among animals whose cerebral hemispheres are well developed, each individual is capable of making at least a partially adequate reply to new environmental demands. Many apparent exceptions to these statements are
to be found, but they are probably only apparent. It is not easy to devise
methods for the measure of cerebral development. This difficulty increases
when a study is made of animals of about the same biological status, or
when an attempt is made to determine a structural foundation for racial
peculiarities, or for variations in the characteristics of different persons
of the same nation or family. It is extremely difficult to find satisfactory
methods for the measurement of the respective degrees of development of
the various parts of the hemispheres.
The phylogenetic development of the cortical centers is rather complex. It is not the purpose of this chapter to give any technical discussion of the relations between the brains of the classes of vertebrates, except as these differences may throw some light upon the development of the structures concerned with the phenomena of consciousness. In the lowest vertebrates, such as amphioxus, there is no brain, in the ordinary sense of the term. The nervous system consists of a tube, with nerve cells and short nerve fibers. The sensory nerve cells are placed upon the roots of the nerves, as in the higher vertebrates, though the migration of the nerve cells in embryonic development suggests the possibility of a primitive type of vertebrate in whom the nerve cells all lie within the walls of the neural tube. In cyclostomes the brain, such as it is, is composed chiefly of the higher olfactory neurons. There is found a layer of cells like epithelium, which forms a sort of anlage of the pallium. Between the amphioxus and the cyclostome are found no intermediate types. The phylogenetic history of the cerebrum thus lacks several chapters at the very beginning. The condition of the thin and non-nervous pallial fold of the cyclostome brain is duplicated in the folding of the walls of the brain vesicles in the development of the choroids plexus. The pallium of the cyclostome is composed only of
a thin layer of non-nervous matter. The brain of this fish consists chiefly
of the olfactory lobes and the connections of the olfactory tracts. The
nuclei associated with the olfactory apparatus in the higher mammals are
found fairly well represented among the cyclostomes. The cortical centers
of olfaction are, of course, not found, since no nerve cells are found
in the pallium. The nervus terminalis is represented slightly in the lamina
terminalis of these brains, but this connection is not found among the
fishes of higher development. It is possible that this relationship of
the nervus terminalis may represent the beginning of a cortical somesthetic
area. Further study is needed in this matter.
In selachians there is found a beginning of cortical structure in the olfactory cortex, or archipallium. This center is closely associated with the centers of the thalamus and striatum, both of which are developed among fishes. In ganoids and bony fishes the olfactory areas alone seem to be functional among the parts of the nervous system comparable to a cerebral cortex. The amphibians and reptiles show a cerebral development which appears to be much greater than is found among the fishes. This might be supposed to underlie a wider cortical representation of the bodily sensations and activities than are possible among the fishes, but this seems not to be the case. The increasing complexity of the reptilian and amphibian brain apparently merely gives opportunity for greater complexity of the relations of the higher olfactory centers. Apparently very suddenly, among lower mammals, there
is to be found the beginning of a neopallium. The archipallium is pushed
toward the lateral and posterior region of the cortex, while the relationships
of the olfactory centers remain practically unchanged.
The entire olfactory nervous apparatus is unique in its retention of its primitive relationships through all the extremely marked variations of the other nerve centers. The neopallium presents a succession of developmental changes during phylogenetic and ontogenetic progress, but the archipalium presents only those variations imposed upon it by the pressure and interposition of other centers and tracts. This conservatism of the olfactory centers, if the rather figurative term may be so used, compares with the physiological relationships of the rhinencephalon among higher animals and mankind. Olfactory centers lie in close relationship to all cortical centers, and these are related very intimately to the basal centers. Thus, it is found in man that, while consciousness of things smelled is not usually very vivid, unless the stimulation is very pronounced, the reactions governed, either directly or indirectly, by the olfactory impulses are decided, involuntary, energetic, and associated with many emotional and instinctive phenomena. The power of the olfactory stimuli to arouse memories of notable vividness is dependent, in part, upon the intimacy of the relationship of the olfactory centers to the centers concerned in the reception and memories of other sensory impulses. In monotremes the olfactory relationships are practically
the same as in reptiles and amphibia. Here the lateral and the posterior
cortical areas are not concerned with the reception and coordination of
olfactory impulses, but begin to assume the place of a somatic pallium.
No reports are available concerning the experimental stimulation of the
cortical centers in these animals, but the structural relations are somewhat
similar to those of the higher mammals. The olfactory centers are even
more highly developed than in non-mammals. The anterior commissure of marsupials
is greatly increased in size, and this is due to the presence of a large
number of the commissural fibers, which afterward assume the position of
the corpus callosum in higher mammals.
The increasing development of the corpus callosum appears to be an important factor in the phylogeny of the mammalian brain. This is associated with a relative diminution of the hippocampus. The primitive hippocampal relations are recalled by the perforating fibers of the corpus callosum. The interposition and increasing development of the corpus callosum pushes aside and distorts the lamina terminalis in a most complex manner. The posterior part of the hippocampus becomes more
and more complex in structure, while the functional relations to other
parts of the nervous system remain practically unchanged.
The hemispheres increase in size much more rapidly
than does the skull, both during ontogenetic and phylogenetic development.
This increase causes the folding which produces the fissures, sulci, convolutions
and gyri of the adult human brain. This increasing complexity of the folded
surface makes possible a great area of cortical structure, which is associated
with increasing complexity of the cell relationships and the cell structure
of the cortex.
The functional development of the cerebral cortex
is not at all well known. Very few experiments have been reported concerning
the effects of stimulation of the various areas of the cortex in the lower
mammals, or of the effects of their experimental ablation. Studies should
be made of the brains of the lower mammals which have been subject to disease
or accident, as well as of those subject to laboratory tests. From our
meager supply of fats, the following general account may be given, with
the understanding that it is, on the whole, very unsatisfactory, and that
it must be subject to considerable modification on further study.
The fishes, amphibia, reptiles, and the marsupials react to the changes in their environment through the intermediation of the lower centers; the midbrain, medullary, thalamic and striate centers are sufficient for the coordination of the movements needful for the proper orientation of the individual to his environmental changes. The olfactory impulses, partly, no doubt, because of the anterior position of the olfactory lobes in the animal body, and partly because of the immense importance of the olfactory impulses in the life history of the animals, are represented by centers of constantly-increasing complexity. Interpolated neurons also increase the possibility of more exact reactions, better adapted to the preservation of the life of the animal, both in seeking food and in avoiding danger. Thus, the olfactory impulses have the honor of being represented in the earliest cortical centers. The nervus terminalis, in the lower fishes, is a
tract which passes from the midbrain centers to the lamina terminalis.
Its specific energy is not known.
Land animals begin to need the sense of hearing and also the sense of touch. Touch, in fishes, must be comparatively useless from the biological standpoint, but among land animals it assumes considerable importance. The touch impulses in these animals are carried by the fiber tracts relating the cerebellar centers to the motor apparatus. By the cerebellar connections the visual and auditory impulses, as well as those arising from variations in temperature and the impulses of touch and pain, initiate the proper reaction to environmental changes. The midbrain and interbrain assume, in part, the functions of coordination in the fishes, as well as in the land animals. The development of the cerebellum among land animals is of interest in this connection. Fish and birds, moving in an element of about the specific gravity of their own bodies, need delicately coordinated neuro-muscular apparatus. They have the cerebellar structures well developed. In the amphibia and reptiles two groups of centers, physiologically distinct, are to be found. The anterior group includes centers which are concerned in the coordination of olfactory impulses, and in facilitating the motor reactions in answer to these. The posterior group includes the centers which are connected in the coordination of all other sensory impulses and the determination of the motor reactions necessary in answer to these impulses. These two groups remain practically distinct, except
for the superseded nervus terminalis in lower fishes, until the mammals
are reached. Then the appearance of the neopallium indicates a beginning
of a cortical representation of the impulses previously coordinated only
by the posterior groups of centers, the midbrain, pons, medulla, cerebellum
and cord. The power of giving variable and complex answers to environmental
demands is not well developed among non-mammals, or among the lower mammals.
In birds the impulses concerned in relating the movements
of the individual to things placed at greater distances become of value
in maintaining life. The anterior part of the midbrain, which is concerned
in coordinating the visual impulses, attains a marked development. The
hemispheres of birds are very small, and there is no reason to believe
that there is any cortical representation of sight. The visual impulses
act through the corpora bigemina of the midbrain, the floor of the aqueduct,
the nuclei of the cranial nerves, the cerebellum, and the spinal centers
by way probably of the tecto-spinal tract. The visual impulses are of great
importance in controlling the movements of the bird, but this does not
indicate whether birds do or do not consciously see and look.
Auditory impulses are of less importance in birds.
The posterior part of the midbrain, which corresponds to the posterior
colliculus, is almost lacking. The vestibular impulses are of considerable
importance in birds, as in fishes, and for the same reason. The bird, as
well as the fish, lives in a medium which has about the same specific gravity
as the body of the individual. The movements must be well governed or a
loss of equilibrium would result. The vestibulo-spinal tracts, as well
as the cerebellar connections of the vestibular nulei, are of great importance
in the proper correlation of the muscular activities which secure motion,
the seeking of food, and the flight, by means of which they are enabled
to resist being themselves used as food.
The sensori-motor areas are not well developed even
in dogs and cats The cruciate sulcus is homologous with the central sulcus,
or fissure of Rolando. The cortex upon the anterior aspect of this sulcus
is concerned in the control of those movements of the skeletal muscles
which are called volitional. The area is related to the lower centers by
the tracts called pyramidal. These tracts are very poorly represented in
dogs, cats, and animals of about that rank. In monkeys these tracts, as
well as the sensori-motor areas, are fairly well represented. In man the
sensori-motor area is of great extent; the impulses from this area are
of great importance in the life of the individual, and the pyramidal tracts
become conspicuous factors in the structure of the spinal cord.
Anterior to the sensori-motor area is found, in man
only, the center for speech. The structure of the cortex of both hemispheres
seems to be adapted to the reception and coordination of the impulses concerned
in the control of the speech movements, but usually only the left speech
center is functional. In persons who use the left hand most, or in whom
some accident or disease has destroyed the left center, the corresponding
group of cells of the right hemisphere may assume the duties of controlling
the speech movements.
With the exception of the olfactory cortex, each cortical sensory area is surrounded almost or quite completely by an overflow area. Between the overflow areas lie areas which are variously called intermediate areas, association areas, or vacant, or silent, or non-functional areas. Of all these terms, the first is most logical. Intermediate areas are those lying between the areas of known function. This term gives no false conceptions concerning the functions or relationships of the areas, and does not pave the way for misunderstandings in the future. In all animals the intermediate areas seem to be
almost or quite non-functional. In mammals of quite a fair development
the ablation of practically the entire cortex, under favorable conditions,
is not followed by any marked loss of nervous functioning. In man practically
all of the left hemisphere and almost all of the right hemisphere are apparently
capable of development. The development of the cells of the cortex and
the progress of the medullation of the cerebral tracts indicate the importance
of the integrity of the cortical centers in civilized mankind. It is true
that most remarkable injuries of brain substance may be suffered without
the appearance of any localizing symptoms. The more highly developed any
person is, however the less is his chance of suffering brain lesions without
symptoms.
The appreciation of the solidity of objects, and of their distance and size, results from the relationship between the visual overflow and the somatic sensory overflow. A center is thus developed in the parietal lobe which is concerned in the correlation of the visual, tactile and muscular images. The extension of areas of developed cells through the overflow areas leads to the structural relationships which underlie the classification of ideas, the consideration of the environmental changes from which sensory impulses arise, and the associational functions by means of which any given reaction may be enabled to make proper reply to any number of factors, though these may have stimulated the nerve centers even years before the reaction is given. The phenomena of the activity of the overflow area are, at times at least, associated with the consciousness of thought, choice, effort, etc., and this is the reason why the unfortunate term psychic has been employed in this connection. Fig. 2. Small and giant pyramidal cells from human
cortex. About 520 diameters. The cells are approximated in drawing, because
the intracellular spaces in the human cortex are so great. The nucleolus
is black, and within the clear nucleus. The granules of yellow pigment
are shown in three of the cells. The dendrites are so long that with this
magnification there would require a cut more than two feet in diameter
to include the dendrites of one cell.
The ontogenetic development of the cerebral cortex is not less complex than the phylogenetic development. Ontogeny repeats phylogeny to a certain extent, though a great many steps of the phylogenetic development are omitted, and other steps are interposed which have no known type in the phylogeny of the nervous system. The entire nervous system arises from the development of cells originally placed upon the surface of the body. In the embryo the epiblast becomes depressed, the cells multiply rapidly; the depression continues until the ridge of cells becomes imbedded in the mesoblastic tissues of the embryo. The epiblast grows over the deep groove, and the cord of cells is thus shut off from the external surface. These cells continue to multiply. Certain of them
divide in such a manner that of the daughter cells one group retains full
power of reproduction, but has little irritability, while another group
of daughter cells retains marked irritability, but has no power of reproduction.
The cells of the first group are called spongioblasts, and from them develop
the neuroglia cells; the other group of cells, neuroblasts, develop into
nerve cells. The actual number of nerve cells of the human body is complete
before the fifth week of embryonic life. After about that period the cells
develop in size, form and function, but they do not increase in numbers.
The neuroglia cells retain the power of reproduction throughout life.
The part of the infolded epiblastic tube which is
to form the brain becomes distended unequally so as to form three vesicles.
The first and the third of these become constricted into two divisions,
thus forming five vesicles. The walls of these five vesicles become developed
into the cerebrum, interbrain, midbrain, pons, medulla and cerebellum.
The walls of the anterior of these vesicles become developed into the prosencephalon,
and it is with the prosencephalon that we are chiefly interested at this
time.
At first a single, rounded vesicle, there soon appears
a depression running antero-posteriorly on the upper aspect of the vesicle.
This depression continues until a deep fold is produced. The infolding
of the wall continues, and the fold is pushed into the cavity of the vesicle,
now apparent as the ventricle. The part of the wall which is pushed into
the ventricular cavity never assumes any nervous characteristics, and is
developed into the epithelial layer of the choroid plexus. Connective tissues
supporting blood vessels push into the ventricles with the cell layer just
mentioned. The choroids plexus of the adult is thus formed.
The lateral walls of the vesicle become developed
into the cortex. The inferior wall becomes greatly thickened, and the cells
become developed into the gray matter of the corpus striatum. The fiber
tracts which push between these cells complete the formation of the striatum.
The optic thalamus is developed in like manner from the walls of the second
vesicle.
At about the fifth week, in the human embryo, there
appears a sulcus or fissure caused by the infolding of the cortex. This
deepens and produces an indentation in the ventricular wall. This fissure
is called the fissure of Sylvius, or the lateral fissure of the cerebrum.
The infolding is continued until a part of the cortex becomes completely
covered from view. The outer fold of the cortex is called the operculum;
the part which is folded within is called the insula, or the island of
Reil.
At a later time during intrauterine and extrauterine
life the infoldings of the cortex continue, and these increase in length,
complexity and depth for a number of years, probably until maturity, and
perhaps longer. It is probable that the development of individual nerve
cells continues throughout life, though no new cells are produced.
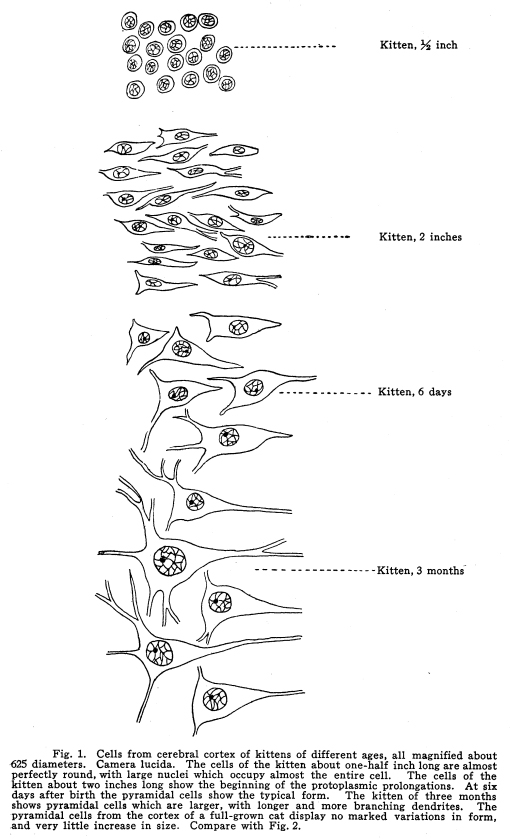
The cells of the nervous system are all alike at the time when the formation of the neuroblasts is complete. The cells then are found, with large, deeply-staining nuclei, and a thin layer of protoplasm of feeble staining power investing the nuclei about equally on all sides. In a ten-weeks human embryo the cells in the different parts of the nervous system are fairly well differentiated The cells of the medullary centers are largest and most highly developed Those of the centers of the midbrain and pons are almost as large. The cells of the spinal cord are becoming arranged in groups, not yet very well defined. The development of the lower part of the cord is less complete than the upper part. The cells of the sensory ganglia can be seen in the intervertebral foramina. The cells of the cerebral cortex are only barely beginning to become differentiated. It is very difficult to distinguish the nerve cells from the neuroglia. Some of the nerve cells show beginning outgrowths, but these are very small, and are not easily distinguished on account of their feeble staining qualities. (Figs. 1, 2.) From the tenth week to the fifth month the change in the size of cortical neurons is less apparent than is the increase in size of the cells of the lower centers. The cord and the medullary, pontine and midbrain centers increase greatly both in size and in peculiarities of outline; but the cortical neurons grow slightly in size, and scarcely more in complexity of outline. Between the fifth month and birth the cortex increases rapidly as a whole and in the size of the cortical cells, in their dendritic and axonic development, and in the assumption of the characteristic staining reactions. Fig. 3. Spindle and polymorphic cells from the external layer of the human cortex. These cells are not frequently found in the brains of lower animals, nor in the brains of of idiots. They are poorly developed in young children, and are most plentiful and best developed in the overflow areas of the adult human cortex. They are also plentifully found in the olfactory cortex. Fig. 4. Cells from the cortex of the rabbits brain.
380 diameters. The irregular appearance of the pyramidal cells and the
short dendrites should be noted.
The histogenesis of the cortical neurons is of a certain interest. The cells first to be differentiated as nerve cells are almost or quite perfectly round. The nucleus occupies almost the entire cell body. A single deeply-staining nucleolus is usually present; rarely there are two or three nucleoli. These nucleoli are, at the first, very much alike. Afterward, as development progresses, one nucleolus seems to be present which is of a slightly different type from the others; indeed, the supernumerary nucleoli seem to be only rather large net-knots in the adult neuron. The network of the nucleus of the embryonic neuron is coarse and stains very deeply. The protoplasm is scanty and stains with such great difficulty that it is often impossible to determine its extent. As development proceeds, the protoplasm displays the most pronounced changes from the embryonic type. After attaining a thickness of about five microns around the nucleus (in the human embryo), the protoplasm begins to branch out in one or more directions. Usually two prolongations are to be found, at opposite sides of the nucleus. This is the case with the sensory cells, with the cells of the higher centers, and, for the most part with the cells of the cortex. In the motor nuclei of the first order, through the whole nervous system, and in the pyramidal cells of the cerebral cortex, there appear at the same time three or more prolongations. The large multipolar cells are sometimes irregular in outline from the first. The more usual form is, however, the bipolar at first, with the various irregularities appearing at a later stage of development. The bipolar form underlies a physiological attribute of the neuron. Throughout life it receives by one process the impulses which affect its metabolism, and discharges by way of another process the nerve impulses which are initiated by its activity. In the case of the sensory neurons of the first order the two processes usually approach one another and finally coalesce for a variable distance from the cell body. The multipolar cells send out many, and long, and freely-branching dendrites, which are probably all receptive in function. The emissive prolongation, called in the adult cell the axon, remains single in almost all cells. In the cerebral cortex the cells, which present at first a perfectly undifferentiated appearance, ultimately become developed into several different types, each with its own particular function. Here are found, first, the peculiar cells, either diaxonic or amacrine, which lie along the surface of the cortex. These receive impulses, probably, from all parts of the nervous system. Among these branch the dendrites of the different layers of pyramidal cells, the collaterals from the axons of the cells of the other layers of the cortex of the same area and perhaps other areas, and the axons of the inverted pyramids. These cells are probably those concerned most immediately in consciousness. They are scarcely to be found in the brains of animals, in any well marked development; they are first injured in certain mental diseases; they are not at all well developed in the brains of idiots or imbeciles, and sometimes are found poorly developed in the feeble-minded The exact place of these superficial cells is not well known. Much more work needs to be done in studying the brains of persons whose mental traits are well known. (Fig. 3.) The cells which develop into the pyramidal cells show their tendencies almost with their first outgrowths. They send one well-marked prolongation toward the surface, and the triangular appearance of the cell, with a single straight, regular prolongation from the center of the base of the triangle, is very characteristic. These cells afterward assume the function of carrying the nerve impulses away from the area where they are found. The largest of the pyramids carry impulses by the longest axons, and, among the very large pyramids of the motor area, there is reason to suppose that the largest cells are those which send impulses which ultimately reach the largest muscles. This relationship is of interest in connection with certain phenomena associated with the symptoms of mania and melancholia. The multipolar and polymorphic cells of the cortex
are developed form the first in an irregular manner. Cells are found in
the lower layers of the embryonic brain (human) which look as if they were
intending to grow into pyramidal cells, but in the adult brain no typical
pyramidal cells are found in the deeper cortical layer.
During the later stages of intrauterine life, and
during the life of the individual, the neurons increase constantly in complexity
of structure. The number of cells resembling embryonic cells becomes progressively
less during early life, and there is some reason to believe that this development
of the individual cells continues throughout the entire lifetime. It is
certain that there is no age limit to the functional development of the
cortical neurons, and the facts of the manner of education among elderly
people, together with the manner of assumption of new duties by cells after
certain injuries or diseases of the brain, points clearly to the supposition
that the functional development is associated with a structural development
of the neurons. In other words, education and mental development are associated
with a structural and functional change in the cortical neurons.
Since there are found large number of neurons of
an embryonic type in the brains of persons who are elderly and whose mentality
is supposed of high order, and since there does not seem to be any limit
to the development of even very old people from the mental standpoint,
it may be concluded that the ontogenetic development of the cerebral cortex
is not complete, even under the most favorable circumstances. It is certainly
true that there is no age limit to the development of the functions of
the cortical neurons.
It seems true, also, that the phylogenetic development of the cortical neurons is not yet complete. There are yet areas upon the surface of the brain, especially in the right hemisphere, whose function either is not known or which have no function at present. Reasoning from the history of the development of civilization, and the development of the mental characteristics of children, it appears now probable that considerable opportunity for increased complexity of associational processesthat is, for increased intellectualityremains even yet upon the cortex. Certainly no modern Alexander need weep for more worlds to conquer if he consider mental attainments of any value. |
{kind=link}
{kind=link}