
|
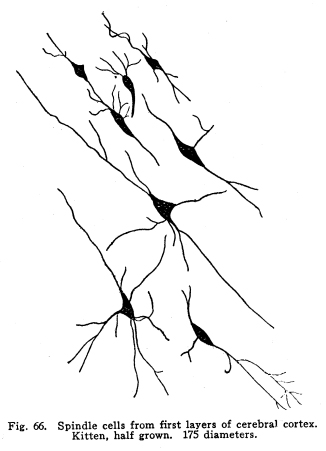
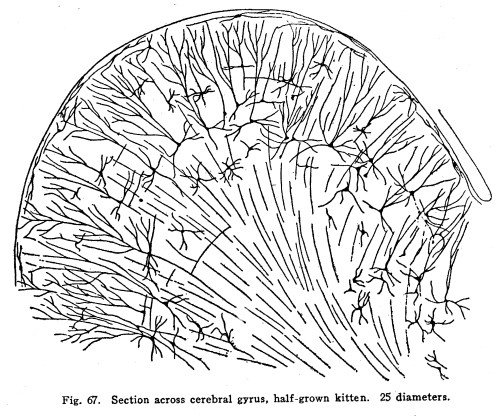
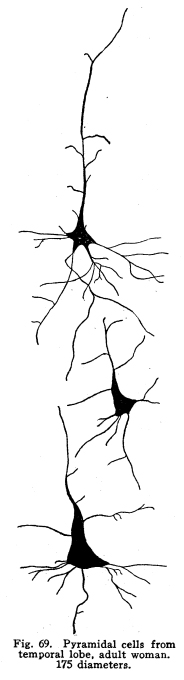
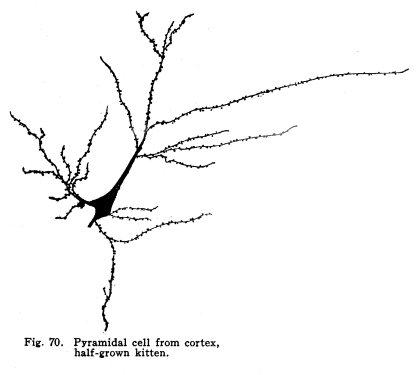
The centers concerned in the immediate control of the various functions of the body have been discussed. It is evident that among these centers there are to be found many variations in complexity. From the simplest reflex actions performed through the agency of the spinal are to the almost infinite complexity of the higher centers there appear all grades of structural variety, with a concomitant functional variety of neuron relationship. But in the cortical centers there is found a degree of complexity, both of structure and of function, which renders the centers already described almost simple by contrast. It is not that the cortical centers present more varieties of structureindeed, the structure of the cortex appears very simple compared with the structure of the olfactory tracts, for examplebut this simple structure is presented upon so broad a plane, and the inter-central relationships are so well developed, that the number of variations in reaction which might follow any given sensory stimulation is beyond imagination. The cerebral hemispheres are covered by a thin and fairly regular layer of gray matter. This layer is called the cerebral cortex. It varies slightly in structure and in thickness in different areas of the cortex, though these variations are not nearly so profound as one would expect when the extreme variations in the functions of the different areas are considered. In thickness there is also some variation. About one-third to one-sixth of an inch is the usual thickness of the cortex. Fig. 66. Spindle cells from first layers of cerebral cortex. Kitten, half grown. 175 diameters. Fig. 67. Section across cerebral gyrus, half-grown kitten. 25 diameters. During embryonic development the nerve cells of the cortex grow with extreme rapidity. The increase in the area of the gray matter is very great, while the associated growth of the white matter, the core of the hemisphere, is comparatively small. As a result of this lack of relative development the gray matter is thrown into a series of folds, called convolutions or sulci. In this way the cortical area becomes comparatively of great extent. The structure of the cortex offers almost inexhaustible facilities for association of neuronic activities. The arrangement of cells and fibers varies somewhat in the different areas of the brain, though the chief factors are everywhere to be found. The classification of the cortical cells into seven layers seems to be accepted. I. The outer layer of cells is covered over and separated from the pia matter only by a layer of neuroglia, which penetrates and supports the nerve cells. These cells are of small size, stellate or spindle-shaped in form, and are either of the amacrine type, of the Golgi Type II, or of similar structure. Among them are found cells which seem to have two or three processes resembling axons. (Figs. 66, 67.) These branch in a T-shaped manner, giving collaterals freely to neighboring cells. This layer of cells receives the incoming fibers from the lower centers and from other areas of the cortex, the collaterals from the axons of the cells of the deeper layers, and the axons of the reversed pyramids. The axons of the cells are all short and do not leave the layer within which they are found. This layer is found more highly developed among races and individuals of high attainments and civilization. It seems that it is the place of most exact coordinations. Fats of clinical evidence indicate that it is in this area that consciousness primarily is affected. The second layer includes the small pyramidal cells, rather closely placed. A few polymorphic cells lie among the pyramids. The apical dendrites of these cells pass toward the periphery of the cortex, and branch freely among the spindle cells of the outer layer. The basal dendrites pass horizontally and branch freely among the other cells of the same layer. The axons of the small pyramidal cells descend to the deeper layers. They are of small diameter, and probably rarely leave the gray matter. These axons give off collaterals, which pass horizontally for a time, giving off branches which form synapses with other cells of the same layer. These collaterals and their branches pass toward the periphery and enter into synaptic relations with the cells of the outer layer. The descending axons of the cells form synapses with the cells of the deeper layers. The third layer resembles the second, and is included with it as the second layer of the older texts. The pyramidal cells are nearly twice the diameter of the small pyramids, and they are pushed further apart by the greater complexity of their dendritic branchings. The fourth layer is the layer of large pyramidal cells. These are of great size, especially in the motor areas of the cortex. Their apical dendrites pass into the outer layers, and branch freely among the cells of these layers. Their basal dendrites branch freely in the same layer. The axons and collaterals of the small and medium pyramidal cells form a network of interlacing dendrites around the bodies and among the dendrites of the large pyramids. The horizontal fibers of these dendrites and axons give the appearance called the line of Baillarger. The axons of the large pyramids enter the white matter. The fifth layer includes the stellate or polymorphic cells in variable numbers. These cells are scarcely to be found in motor areas, but are well marked in the auditory and visual areas. They are found in the post-central convolutions. The dendrites of these cells branch freely, but are not very long. Their axons pass horizontally within the same layer. Either the axons themselves or their collaterals pass to the outer cell layer to branch among the stellate and spindle-shaped cells. Fig. 68. Pyramidal cells, occipital lobe, new born baby. 175 diameters. The sixth layer is the inner layer of large pyramids. In the motor areas these are of very great size. It includes many cells of the Golgi Type II, as well as polymorphic cells. The giant cells of this layer are characteristic of the motor area. These, as well as the large pyramids of the fourth layer, are found degenerated in amyotrophic lateral sclerosis. Their place as the origin of the pyramidal tracts is thus demonstrated. (Figs. 68, 69, 70.) All of the large pyramidal cells contain great masses of tigroid substance (Nissl), and these masses are found absent after long-continued overwork of the cells, as after epileptic attacks or convulsions. The seventh layer includes polymorphic cells, chiefly of the spindle variety. (Figs. 71, 72.) The cells have several dendrites, which are often very long and may or may not branch very freely. Their axons enter the white matter and pass to adjacent or distant areas of the cortex. Some pass toward the outer layer of the cortex. This layer includes many cells of the Golgi Type II and a few inverted pyramids. The axons of the latter pass into the outer layer of the cortex, giving collaterals to all layers in passing. Fig. 69. Pyramidal cells from temporal lobe, adult woman. 175 diameters. Fig. 70. Pyrramidal cell from cortex, half-grown kitten. Fig. 71. Polymorphic cells from seventh layer of occipital lobe, new-born baby. 190 diameters. Fig. 72. Hippocampus, half-grown kitten. 10 diameters. The different areas of the cerebral cortex are occupied by centers for the performance of different functions. The factors concerned in the primary localizations are not well understood. In the cortex of lower mammals very little differentiation of structure is to be found. With increasing development of any cortical function the centers concerned in that function show increasing complexity and greater peculiarities of structure. It seems that the assumption of any function by any cortical area is associated with such a change in the structure of that area as best fits it for the performance of those duties. It is apparent that the assumption of any particular duty by any particular cortical center must be due to one or several conditions. The certain area most accessible to the sensory impulses concerned in the control of any specific function would, other things being equal, assume most readily those duties. On the other hand, the areas of the cortex in the lower mammals, while they appear alike throughout, may be found to possess variations which render them more fit for the performance of one duty than another. The fact that among the various classes of mammals the same arrangement of centers upon the cortex is presented is evidence that the structural adaptations to the performance of the different functions are an essential part of the lower mammalian, and probably also of the vertebrate, morphology. In the development of the cortical centers the relation between structure and function is shown. The structure seems to be essential to the assumption of duties; the assumption of the duties leads to the development of the structure better adapted to the performance of those duties. The performance of function and the development of structure occur together. Both together make the greater efficiency, which is the reply of living creatures to the environmental demands. The areas which are recognized as possessing specific functions do not include the entire cortex. Each specific function is surrounded by an area of cells which, in the lower mammals, seem to have no function, and in man are not known to have specific functions. There are large expanses of cortex which appear to contain no centers and to be non-functional. The indifferent areas are of less extent in mammals of higher development. The area surrounding the center for any specific function has been called the psychic center for that function. This term is very unfortunate. In the first place, the term psychic gives the impression of consciousness. It is not known that there is any more mentality or consciousness concerned in the activity of the psychic areas than in the activity of the other cortical areas. It is unfortunate to employ words of philosophical significance to physiological phenomena. The word psychic has its own significance; it is already a useful term, and to apply it to a structural area adds unnecessary confusion and lessens its proper value. For these reasons I have ventured to employ the term overflow in this connection. The overflow of any center is the adjacent cortical area whose cells are subject to stimulation by that center. No psychological significance is attached to the word, and it must be remembered that the term refers to the structural relationship only. It is yet to be determined what relationship exists between the activity of the cortical areas and the phenomena of consciousness. The
cortical centers are most exactly explained by considering the sensory
centers or areas first.
The common sensory area includes the posterior wall of the central sulcus, the anterior half of the posterior central gyrus (ascending parietal convolution), and the adjacent part of the paracentral lobule. This area is divided into smaller areas, which are the centers of the different bodily areas. All epicritic and a part of the protopathic sensations are consciously appreciated by means of the activity of these areas. Disease of this area causes loss of sensation of those parts of the body in relationship with the affected centers. In locomotor ataxia the area is found degenerated after the method of Nissl. The centers for common sensation receive impulses as follows: I. Epicritic sensations are carried from the lower part of the body and the lower limbs by way of the sensory nerves, sensory ganglia, posterior roots, fasciculuc gracilis, nucleus gracilis, the median fillet (decussating), the lateral nucleus of the thalamus, the thalamic radiations to the cerebral cortex. Certain of the impulses of this class seem to be carried as described to the nucleus gracilis, from this point by way of the restiform body to the cortex of the superior vermis of the cerebellum, thence to the nucleus dentatus, then by way of the brachium conjunctivum to the red nucleus, then to the optic thalamus, probably the globus pallidus, and the cerebral cortex. II. Epicritic sensations are carried from the upper limbs, the upper part of the body, including the neck, as high as the lower line of the mandible, by way of the sensory nerves and ganglia, the posterior roots, the fasciculus cuneatus, the nucleus cuneatus, the median fillet, the lateral nucleus of the thalamus, the thalamic radiations and the cortex of the opposite side. The crossing occurs at the decussation of the fillet, in the medulla. III. Protopathic sensations from the entire body, and probably some of the visceral sensations, are carried by way of the sensory nerves and ganglia, the posterior roots, the cells of the dorsal nucleus or the posterior horn, thence to the opposite side into the spino-thalamic tract, to the lateral nucleus of the thalamus, and to the cortex. IV. Visceral and perhaps also protopathic sensations are carried by way of the sensory nerves and ganglia, the posterior roots, the cells of the dorsal nucleus (Clarkes column), thence to the lateral ascending cerebellar tract of the same side, and through the restiform body to the cortex of the superior vermis of the cerebellum; from the cerebellar cortex the impulses are carried to the nucleus dentatus, thence by the brachium conjunctivum to the red nucleus, thence to the thalamus, the caudate nucleus, and the cerebral cortex. V. Similar impulses are carried by the same path to the dorsal nuleus of the cord, thence by way of the ascending cerebellar tract and the brachium conjunctivum to the cortex of the superior vermis of the cerebellum, then, as in the case of IV, to the cerebral cortex. VI. The cranial nerves of common sensation send impulses by way of their nuclei of insertion to the cerebellum, and thence by the pathway already described to the cerebral cortex, and they send impulses by way of the median fillet to the lateral nucleus of the thalamus, and thence to the cerebral cortex by the thalamic radiations. VII. Impulses from corresponding areas of the opposite side of the brain reach each area of common sensation. VIII. Impulses from adjacent cortical areas, and from distant cortical areas, reach the areas for common sensation. The area for common sensation sends impulses as follows: I. Impulses are carried by short fibers to the sensory overflow, in which probably memories of the body sensations are stored. II. Impulses pass to the motor areas in the anterior wall of the central sulcus, and in the adjacent part of the paracentral lobule. III. Impulses pass to the lower centers, the thalamus, the corpora striata, the smaller basal ganglia. IV. Impulses pass to corresponding areas in the opposite hemisphere, and to other parts of the same hemisphere, but chiefly to the frontal lobes. This
area, then, receives all common sensations from the body, the epicritic
sensations by a pathway of direct and with few relay stations, the protopathic
and visceral by a longer, indirect pathway, with many interposed neuron
systems. The impulses thus received are transmitted to the basal
ganglia, and are concerned in modifying the instinctive reactions, and
to the frontal lobes, especially the left, in which the impulses concerned
in the ideas of personality, of ones own position in the midst of his
environment, are coordinated.
The area concerned in hearing is place din the first and second temporal gyri. This area receives impulses as follows: I. Impulses aroused by the stimulation of the hair cells of the organ of Corti in the middle ear are carried by the dendrites of the auditory cells, the ganglion spirale, the cochlear portion of the auditory nerve to the nuclei of termination of the auditory nerve (the lateral and the median nuclei), thence by way of the trapezoid body, decussating, into the lateral fillet; after an uncertain number of relays in the nuleus of the trapezoid body, the superior olivary nucleus and the accessory nuclei of the superior olivary body, the impulses are carried by the lateral fillet to the inferior colliculus, thence by the inferior brachium, and also by way of the fillet, to the internal geniculate body, the acustic radiations, and the cortex of the first and second gyri of the temporal lobe. II. The areas for hearing receive impulses from corresponding areas of the opposite hemisphere; from neighboring and distant areas of the same hemisphere. The acustic area, or center, sends impulses as follows: I. By way of the acustic radiations, the internal geniculate body, the inferior brachium and the inferior quadrigeminates, the descending fibers of the lateral fillet, thence by the nuclei of insertion of the auditory nerves the nuclei of the motor nerves for the auditory muscles, the vaso-motor centers for the internal ear, to the cochlea and the structures directly or indirectly concerned in hearing. II. The impulses initiated by the activity of the cells of the acustic area are carried by short association fibers to the auditory overflow, in which are probably located the areas for associative memories of things heard. III.
Impulses are carried by long association tracts to the distant areas of
the cortex, of the same hemisphere, and to the corresponding areas of the
opposite hemisphere.
The cortical center for the consciousness of seeing occupies the cuneus and lingual gyrus. The visual overflow includes a rather extensive neighboring area of the cortex; the exact limits of this area are not well known at present. The cuneus and the lingual gyrus seem to receive the impulses initiated in the retina, and consciousness is affected by the functional activity of the cortical cells of this area. The stimulation of these cells causes the stimulation of the neighboring cells of the visual overflow, and the activity of these cells is probably concerned in the appreciation of the significance of things seen. The visual center receives impulses from the following sources: I. The visual center of each hemisphere receives the impulses initiated in the yellow spot of the retinae of both eyes. II. The impulses initiated in the homolateral halves of both retinae reach the visual center. These impulses, as well as those of I, are carried as follows: The light affects first the rods and cones. The stimulation of these causes the stimulation of the bipolar cells of the retina, which in turn stimulate the ganglionic cells of the retina. The axons of the ganglionic cells make up the optic nerves. These pass backward to the optic chiasma. Here the nasal halves of both retinae decussate, while the temporal halves persist upon the same side. The fibers, the same axons of the ganglionic cells, are called optic tracts porterior to the chiasma. The term tract properly belongs to the bundles of axons of the ganglionic cells through their entire extent. These tracts give off a bundle which passes to the anterior quadrigeminates, in which we have at present no interest. The most of the fibers of the optic tract enter the lateral geniculate body and the pulvinar of the thalamus. The latter represents the higher development. There are certain relay stations in the geniculate body and the thalamus, no one can tell how many, and the relationship between the thalamus and geniculate body is very intimate. The cells of these bodies send axons in the optic radiations to the cortex of the cuneus and the lingual gyrus. III. The optic center receives impulses from the corresponding areas of the opposite hemisphere, and from practically all parts of the same hemisphere. The optic center sends out impulses as follows: I. Impulses pass downward, following the ascending path of the sensory impulses, by way of the optic radiations, the pulvinar and geniculate body, the optic tracts, to the retinae. These impulses are apparently concerned in some way with the nutrition of the retinae. Impulses from the geniculate body, originating in the cortex as in the case just mentioned, pass to the anterior quadrigeminate by way of the superior brachium. Probably the coordination of the eye structures in answer to the needs of conscious seeing are thus secured. II. Impulses pass by short association fibers into the visual overflow. III. Impulses pass by long association fibers to the opposite visual center, and to other parts of the same hemisphere. IV
.Impulses are carried to the central ganglia. The relationship of
these fibers has not been sufficiently studied.
The cortical areas for taste have not been outlined with any degree of certainty. The few facts which are known in this connection seem to indicate that taste and smell have cortical areas which are very closely related, if not, indeed, actually identical. For this reason the olfactory areas, which are fairly well known, will be described, and the gustatory left with the rather unsatisfactory statement that the description of one probably applies, with at least a fair degree of accuracy, to the other. The uncus hippocampi probably is to be considered the chief center for smell. Either the fusiform gyrus or the cingulum may be considered the center for taste, though all of these areas are included in the rhinencephalon. Since the olfactory impulses secured the very earliest cortical representation, they are to be held worthy the respect due to ancient lineage. Phylogenetically the oldest, the olfactory centers have been subjected to much pressure from the developing centers for other functions. The olfactory areas have been pushed aside, and distorted, and compelled to new connections, and to constant rearrangements in order that they might remain adapted to the changing conditions caused by the constant and almost unending processes of cephalization. In this way the centers concerned in smell and to a certain extent in taste have become of so great structural complexity. The olfactory centers have been described already in a previous chapter. The connections of the uncus and the cingulum may be very briefly given. The olfactory and gustatory areas receive impulses from the following centers: I. Impulses from the olfactory membrane pass by way of the olfactory nerves, the olfactory glomeruli, the mitral cells, and their axons as they make up the olfactory tracts to the olfactory areas at the base of the brain. Here the impulses are carried by three or four different roots to the uncus hippocampi, the inferior gyrus of the temporal lobe, the amygdaloid nucleus, and various parts of the limbic lobe. The uncus seems to be the chief center for consciousness of smell. II. Impulses concerned in taste are carried by way of the sensory nuclei of the fifth, seventh and ninth cranial nerves, the median fillet, the lateral nucleus of the thalamus, perhaps the caudate nucleus and the globus pallidus, to the taste centers, probably in the gyrus cinguli or the fusiform gyrus, with other connections in the neighboring parts of the rhinencephalon. III .Impulses of touch, temperature, and probably of pain are transmitted from the somesthetic areas to the centers for both taste and smell. In this manner the taste and the smell are both subject to modification by the other sensory impulses. It is also true that the temperature and touch changes modify the circulation and other local conditions of the receptive membranes. IV. All parts of the rhinencephalon are intimately associated by small nerve tracts of association. V .The olfactory area receives association fibers from nearly all of the lower centers of the cerebrum. The relationship which the sense of smell bears to the instinctive and emotional reactions is thus, in part, secured. VI .The olfactory area receives very few association fibers from other parts of the cortex. The relationship between the smell of things and the sight or the sounds of things is thus rather scanty. The olfactory area sends impulses to other centers as follows: I. Impulses from the olfactory area are carried to the frontal lobes, and to a less extent to the parietal lobes, by association tracts. II. Impulses pass downward, following the ascending path, to be distributed finally to the olfactory lobes, and perhaps the impulses thus carried may be transmitted by descending fibers to the olfactory membrane. III. Impulses pass to the nucleus habenulae, from which they are carried by the fasciculus retroflexus (Meynerts) to the interpeduncular ganglion, and probably to the motor nuclei of the cranial and certain spinal motor nerves. Thus the olfactory impulses are able to initiate skeletal movements. IV. Impulses are carried by way of the fornix to the corpora mammillaria. From these bodies fibers pass to the thalamus, and to the gray matter around the cerebral aqueduct. The latter pathway transmits the impulses to the nuclei of the viscero-motor cranial, and perhaps certain spinal nerves. Thus impulses of smell are able to modify visceral activities. V. Impulses from the different parts of the rhinencephalon are carried by many small nerve tracts to practically all of the nuclei of the corpora striata, the thalamus, the sub and hypothalamic centers, and the substantia nigra, red nucleus, etc. In this way the impulses aroused through odors are able to affect the emotional and instinctive actions. VI.
Impulses from the rhinencephalon are carried to the other parts of the
cortex. In this way the information brought through the olfactory
tracts may be correlated with the information brought by other sensory
tracts. This relationship is much less exact, so far as consciousness
is concerned, than are the relationships between the other senses.
On the other hand, the impulses aroused by odors, by the very fact of their
poor relationships, are in themselves very efficient in arousing efferent
impulses.
The center concerned in the appreciation of things as having three dimensions, the stereognostic center, lies in the superior parietal lobule and the precuneus. This area is continuous, posteriorly, with the visual overflow, and anteriorly with the somesthetic overflow. The stereognostic image is primarily a function of touch, and in part also of muscular effort. The application of this knowledge to the interpretation of visual images affords a very important addition to the value of sight in adding to the knowledge the individual has of his more extended environment. The stereognostic center receives impulses as follows: I. Chiefly, the center is affected by impulses from the somesthetic centers. II. An important stream of impulses is carried from the visual overflow. III. Impulses from the opposite hemisphere are transmitted to this area, and impulses from other centers of the same hemisphere are sent to the stereognostic center. Impulses from the cells of the stereognostic center are carried to the following destinations: I. Impulses are sent to the motor centers. The efficiency of the volitional coordinations is thus increased. II. Impulses are sent to the visual, and, to a slight extent, to the auditory, overflow. The appreciation of space and of extension in three directions is thus added to the psychical content of the auditory and visual images. III. Impulses are sent to other parts of the cortex, and to the homologous center of the opposite hemisphere. Only a few records are found of cases in which lesion of this center has been associated with localizing symptoms before death. Unfortunately, the capability to describe vividly the symptoms resulting from lesion in this area requires a certain degree of intelligence. The ignorant person, suffering such a lesion, would find himself unable to describe intelligibly his sensations. In
the very few cases whose record I am able to find, the lesion has been
associated with a peculiar sensation of unreality. The patient loses
the power of appreciating the three dimensions of space; he loses his sense
of the solidity of objects, and thus his sense of reality. His attitude
toward the third dimension of space is similar to the mental attitude of
the normal person to the fourth dimension; it is simply unthinkable.
Such a patient, losing his sense of solidity, or reality, becomes unable
to realize the existence of objects beyond his immediate environment.
So far as his perceptions are concerned, the very fact of his turning his
attention away from any object blots it immediately from existence.
He becomes unable to perceive objects as having actual existence apart
from their immediate and transitory effects upon himself. His mental
attitude to any object apart from his immediate environment is similar
to the attitude of the normal person to a perfectly remembered dream.
There is no question of loss of memory; the actual existence of the thing
seen in a dream is totally outside of the range of recognized possibilityoften
is itself utterly unthinkable. Yet one may remember his dream perfectly,
and the patient with the lesion in the stereognostic area may remember
the things seen and felt, while finding their continued existence apart
from their effects upon his sense likewise unthinkable. Astereognosis
is, then, comparable to mind blindness, or mind deafness.
The motor area includes a number of centers lying along the anterior portion of the postcentral gyrus (ascending frontal convolution), and in the neighboring area of the paracentral lobule. The motor overflow extends toward the front of the area mentioned. In the motor overflow the movement memories are probably located, and the centers concerned in the control of the immediate motor neurons. Probably in the motor overflow are found centers whose activity coordinates the activities of the cells of the motor area, much as the association cells of the cord coordinate the spinal reflexes. The motor center receives impulses as follows: I. Impulses from the somesthetic area of the same side, possibly of the opposite side, may affect the activity of the motor cells. This reaction may or may not be conscious. II. Impulses from the association centers may cause increased or decreased action of the motor cells. III
.Impulses from the speech center affect the activities of the cells motor
to the voice muscles.
The motor center sends impulses as follows: I. Chiefly, the fibers from the large pyramidal cells of the motor area descend into the internal capsule, through the crura, the pons and medulla, to become the pyramidal tracts. These fibers ultimately terminate in the center of the gray crescent in the spinal cord and around the cells of the motor nuclei of the cranial nerves. These fibers are carried to the somatic motor, but not to the visceral motor nuclei, under normal conditions. II. Axons and collaterals from the pyramidal tracts terminate in the optic thalamus, the corpora striata, the substantia nigra, the red nucleus, and probably other of the basal ganglia and related centers, and into the corpus callosum. III Axons from cells of the motor area, probably of all layers, carry associational impulses to all other cortical areas. Lesion
of any motor center is associated with paralysis of the upper neuron type
of the muscles controlled by that center.
This area seems to be developed, in right-handed persons, to a greater extent in the left side of the brain than on the right. The anterior association area is continuous, on its posterior aspect, with the somesthetic area. It seems probable, from a study of the phylogenetic development of the frontal lobes, that whatever sensory impulses from the viscera reach the cortical centers at all are carried to the frontal lobes. These areas are separated from the other sense areas of the brain, except as they are associated through the long association tracts. The anterior association area is, then, admirably adapted, structurally, to the coordination of those impulses concerned in the consciousness of personality, the appreciation of external changes in the light of their effects upon self. This area receives impulses from the following sources: I. Impulses are carried from the viscera, by way of the indirect sensory paths through the cerebellum and the spino-thalamic tracts, and by way of the vagus, to the thalamus by the fillet, and to the cortex by the thalamic radiations. II. Impulses are probably carried by way of the spinal sensory tracts, as in the case of the somesthetic areas, directly to the frontal lobes. III .Impulses from the somesthetic areas of the same side of the cortex are carried to the frontal lobes. IV .Impulses from the homologous areas of the opposite hemisphere are carried by way of the corpus callosum, probably in part by indirect paths through the commissures. V. Long association tracts carry impulses from every other part of the cortex to the frontal lobes, especially of the left side. The anterior association area sends impulses as follows: I. The anterior association area sends impulses, partly if not chiefly inhibitor, to the various centers at the base of the hemispheres. II. This area sends impulses to other parts of the hemisphere of the same side, and to the homologous area of the opposite hemisphere. III. Impulses concerned in the production of volitional movements are carried to the motor center. IV. Impulses concerned in the performance of the more delicate and highly coordinated acts probably are carried by way of the red nucleus and the brachium conjunctivum to the cerebellum without much, if any, associated stimulation of the motor area. V. Impulses from the anterior to the middle and the posterior association areas are concerned in the coordination of the ideas of complex relationships involving the place of personality and of many environmental factors. Injuries
or disease of the frontal lobes of the left side, exclusive of the speech
center, are associated with symptoms of uncontrolled emotions and instinctive
reactions (see I, above), and with a loss of the patients place in the
midst of things, of his relation to his neighbors and his environment.
Rarely the lesions of the frontal lobe alone are associated with stupidity
and lack of emotional actions, unless there is also a condition of greatly-increased
intracranial pressure.
This is the insula, or island, of the cerebrum (Island of Reil). It lies beneath the cortex, being formed by the infolding of the cortex, which is associated with the formation of the lateral fissure of the cerebrum (fissure of Sylvius). The cortex of the middle association area is continuous with the speech center, the auditory overlap, the motor center, the somesthetic center, and to a slight extent with the visual overlap. Its
function is not well known. Lesions in this area have been associated
with paraphasia, with visual aphasia, and sometimes with no recognizable
localizing symptoms before death. The connections of the center are
not known, beyond the fact that fibers pass to and from almost or quite
all of the other association areas, and probably also other cerebral centers.
This area also is most highly developed upon the left side of the brain in the ordinary right-handed person. This area is intimately associated with the centers of vision and the visual overflow, with the centers for hearing and the auditory overflow, with the centers for somatic sensations, and with the common sensory overflow. For this reason the posterior association area is so placed as to be adapted to the coordination of these impulses arising from the incoming sensations both general and special, and from the appreciation of the significance of these sensations, as they are received and modified and held through the agency of the neighboring psychical or overflow areas. The posterior association areas receive impulses from the following sources: I. From the visual, auditory, common sensory and their neighboring areas, both by short and by long association tracts. II. From the anterior and the middle association areas. III. From homologous centers on the opposite hemisphere. IV. From the corpora striata and the optic thalamus, and perhaps from other centers associated with these. The posterior association area sends impulses to other centers, as follows: I. Impulses concerned in the performance of movements are sent to the motor area. II. Impulses concerned in speech coordinations are sent to the speech center and the middle association area. III. Impulses concerned in the performance of habitual and delicately coordinated acts are probably sent by way of the red nucleus, the brachium conjunctivum and the cerebellum. IV. Impulses are sent to the middle and anterior association areas which are concerned in the transmission and coordination of the impulses concerned in the more logical mental actsthose underlying the manifestations of the higher intelligence. Injuries
or disease of the posterior association area of the left side are usually
accompanied by a loss of the higher mental faculties. Loss of control
and the performance of improper emotional and instinctive acts are not
characteristic of lesions of this part of the brain. The patient
loses the power to concentrate his thoughts, to work out problems requiring
logic or classification or the power of anticipating events; but he is
usually docile, self-controlled, and his faults are those rather of stupidity
than of passion. It may happen that other symptoms are found in special
cases.
The centers concerned in the expression and recognition of thought are practically limited to mankind. Among animals, the expression of feelings, and, to a certain extent, the conveyance of information of environmental changes, is to be found, though in a very minor degree, when compared with the capacity of mankind. It is not known that thought is to be found among animals as a conscious phenomena, though it is a matter of common knowledge that animals sometimes act as if they had thought. The language centers probably include in their metabolism and structure some of the most complex and highly specialized activities of which nervous matter is capable. The centers for language include a number of different areas, each of which has its own function. Of these centers, it is probable that each is associated with every other by fiber tracts, and that all of these receive association fibers from the different association areas, from the basal ganglia, from the visual and the auditory overflows, and perhaps from the rhinencephalon, either directly or indirectly. The impulses from each center probably are carried, either directly or indirectly, to the centers of the muscles for the control concerned in the actions necessary to the expression of the thought in some particular manner. The impulses from the writing center, for example, must ultimately reach the motor area for the arm and finger muscles. The language centers include the following: The motor speech center occupies the cortex of the left third frontal gyrus. Injury of this center loses the power of speech, though there is no paralysis of the muscles concerned in speech. The patient is unable to form in his imagination the positions of the vocal organs; the muscular memories of speech-making are lost. The writing center occupies the second frontal gyrus. Lesion of this area causes the loss of the power to write, though no paralysis of the arm or finger muscles is present, nor is there any loss of mentality. The center for the recognition of the significance of words, the word-naming center, occupies the angular gyrus. The injury of this center causes the loss of the power to write, because that which might be written cannot be recognized. Lesion of this center causes loss of the power to read. Words are seen, their forms may be remembered, but the significance of the letters and the words they form are not to be recognized. The condition is that of word blindness. A center for the recognition of the significance of objects has been described in the right angular gyrus. Injury of this center causes the loss of the memories of the use of objects. Such a patient should be able to see perfectly, and even to name an object, but he is not able to remember its appearance, nor to determine its use or significance. A center for gesticulation has been described as occupying the region bordering upon the motor centers for the muscles employed in gesticulations. It seems that such a center must be rather intimately associated with the lower centers for the expression of the instinctive and emotional acts. The evidence in favor of a cortical localization for gesticulation is not conclusive. The auditory word center occupies the posterior portion of the first temporal gyrus. Lesion of this area causes word deafness. It is usually associated with some motor aphasia. A center for tone deafness has been described in the second temporal gyrus. These centers are probably complex in their structure. Probably each center includes a variable number of subordinate centers. The loss of the use of nouns, for example, or of certain languages in polyglots, the phenomena of paraphasia, all indicate that the physiology of the language centers is more complicated than at first appears. |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}