
|
The midbrain is derived from the middle cerebral vesicle. It is proportionately less well developed in the higher vertebrates, especially the higher mammals, because of the enormously greater development of the cerebrum. The midbrain is, however, actually better developed, in certain senses, in the higher mammals, and especially in man, since it is here that we find the greater differentiation of the mesencephalic cells and the greater specialization of the midbrain functions. The midbrain in man is about a half inch in the antero-posterior direction, rather more than an inch in vertical diameter, and from an inch to one and a half inches transversely. It is traversed by a channel, the cerebral aqueduct, or the aqueduct of the cerebrum, or the aqueduct of Sylvius, or the iter a tertio ad quartum ventriculum, as it is variously called. The aqueduct is about a half inch in length. It is continuous anteriorly with the third ventricle, and posteriorly with the fourth ventricle. It is lined with columnar epithelium, which is ciliated during embryonic life in all vertebrates studied, and the cilia may be found throughout life in some animals. The posterior aspect of the midbrain is occupied by the quadrigeminate bodies. The superior colliculus is composed of the anterior pair of the quadrigeminates, which are sometimes called the nates. The inferior colliculus is composed of the posterior pair, which are sometimes called the testes. The superior surface of the midbrain is continuous with the thalamus and the bases pedunculi. The anterior or ventral aspect presents a deep median sulcus, on either side of which lie the convex surfaces of the peduncles of the cerebrum. External to the curving surface the shallow oculo-motor groove shows the superficial origin of the third cranial nerve. The
inferior surface is continuous with the pons and the superior brachium.
The quadrigeminate region shows considerable changes in its phylogenetic development. In the lower fishes the midbrain is composed of a simple tube corresponding to two segments or neuromeres. With the progress of cephalization the thickening of the roof of these segments gives place for the termination of certain of the sensory tractsthe axons of the sensory neurons of the second order from the somatic and visceral sensory nuclei of the medulla and pons, the nucleus gracilis and nucleus cuneatus and the sensory nuclei of the fifth, seventh, eighth, ninth and tenth cranial nerves, and the axons of the second or third orders from the retina. In lower fishes these sensory terminations seem confused and intermingled inextricably. With the increasing development of the quadrigeminates the roof of the midbrain broadens and the aqueduct is proportionately widened, so that it approaches the size of the other ventricles. The quadrigeminates in the lower fishes seem to represent the place of most exact coordination of the nerve impulses concerned in adapting the bodily movements to the environmental changes. The efferent impulses are carried by paths similar to those of higher animals to the somatic muscles of the entire body. In the amphibia the lateral line sense organs are not found, the eyes are of comparatively less biological value than in fishes, and there is less need for delicately coordinated muscular movements on land than in water. The quadrigeminal region is found very much less developed in amphibia for this reason. The increase in the cephalization begins to over-shadow the quadrigeminates, also, and a beginning of the accumulation of the cells concerned in the auditory impulses in the posterior portion and of the cells concerned in visual impulses in the anterior portion is found in amphibia. The aqueduct regains its small size. In birds another change is found. Birds find visual impulses of great value, and their muscular movements must be subject to delicately coordinated nerve impulses. The midbrain in birds attains considerable development, both in the connections of the neurons concerned in vision and also in the connection with the lower centers. The tecto-spinal tract in the anterior longitudinal bundle is here, as in fishes, of great value in securing the muscular coordinations necessary to the maintenance of equilibrium. The birds show no division of anterior and posterior colliculi, partly because the differentiation of the midbrain roof nuclei is not complete, and partly because of the comparatively less development of the auditory centers, and the less important biological value of the auditory impulses. In
mammals the specialization of the quadrigeminal region is fairly complete.
The development of the higher centers leaves the sensory supply of the
quadrigeminates comparatively scanty, while the retention of certain sensory
connections with these cells, whose axons yet pass toward certain motor
nuclei, has led them to assume the less general and more highly specialized
functions concerned in the coordination of the eye and ear movements, nutrition
and control, and in the control of those other structures whose activity
affects the eye and ear tissues or functions, or whose activity the auditory
and visual impulses may affect.
The superior colliculus is composed of two bodies, one on either side. These bodies present a more complex structure than do those of the inferior colliculus. The superior collilculus receives about one-fifth of the fibers of the optic tract. These fibers circle around the body, making its external layer of fibers. A layer of neuroglia invests the outer aspect of this layer. These fibers enter the mass of the body, and form synapses with the cells of the deeper layers. The more superficial cell layer is composed of cells somewhat resembling those of the cerebral cortex. They are small, polygonal and multipolar, sometimes spindle-shaped. This layer is separated from the lower layer of cells by white fibers, partly those of the optic tract, partly those from the occipital cortex, and partly other unidentified fiber tracts. The deeper cells are larger, multipolar and pyramidal, and have very long and complexly dividing dendrites. The structure of this body is described as being composed of six layers: (1) The neuroglia layer, (2)optic fiber layer, (3) layer of small nerve cells, (4) inner medullated layer, (5) layer of large nerve cells, (6) central gray matter around the aqueduct. The superior colliculus receives impulses from the following sources: I. The optic tract is, in mammals and man, the most important source of impulses. II. A few fibers from the medial fillet enter the body. This represents an important original relationship, now superseded to a great degree. III. Fibers from the thalamus and other neighboring centers bring the activity of all into normal relationship. IV. Descending fibers from the cortex, chiefly of the occipital lobes of the same side, enter the body. The impulses carried by these fibers are concerned in the nutrition of the eyeball and related tissues, in some manner not understood. V.
Ascending fibers from the cerebellum by way of the brachium conjunctivum
reach the body. These fibers probably carry two classes of impulses:
First, the indirect visceral sensory impulses are carried to the body,
partly as a remnant of the primeval relationship, and these are manifest
in the injury to the functions of the eye produced in certain forms of
visceral disturbance. A second class of fibers carried from the cerebellum
to the superior colliculus carry the impulses from the cerebellum, which
bring the eye movements into coordinate relationship with the other somatic
muscles.
I. Fibers, axons of the cells of the superior colliculus, probably both of the large and the small cells, pass around the aqueduct to end about the cells of the viscero-motor and the somato-motor nuclei of the third cranial nerve, and of the somato-motor nuclei of the fourth and sixth cranial nerves. In this way the direct reflexes of the eye are coordinated. II. Impulses are carried, either directly or through the intermediation of other neurons, to the pupilo-dilator center in the pons. III. Impulses are carried to the vaso-motor center in the lower part of the medulla. These are concerned in the control of the circulation of the orbital structures. IV. Impulses are carried by way of the tecto-spinal tract in the anterior longitudinal bundle to the cilio-spinal center in the upper thoracic segments of the cord. V. Impulses are carried by the same pathway to the segments of the cord through its whole extent. These are reminiscent relations; and while the impulses so carried are probably of a certain value in the maintenance of muscular tone, it is not probable that they are of any great importance. In diseases affecting the quadrigeminates no loss of muscular tone is perceptible. VI. Efferent fibers from the superior quadrigeminates pass with the optic nerve to the retina. In the injury of these fibers the nutrition of the retinal tissues, and, indeed, of the eyeball generally, is rendered abnormal. VII.
Fibers from the body are carried to other neighboring gray masses, partly
as reminiscent structures and partly as a means of securing coordinate
action.
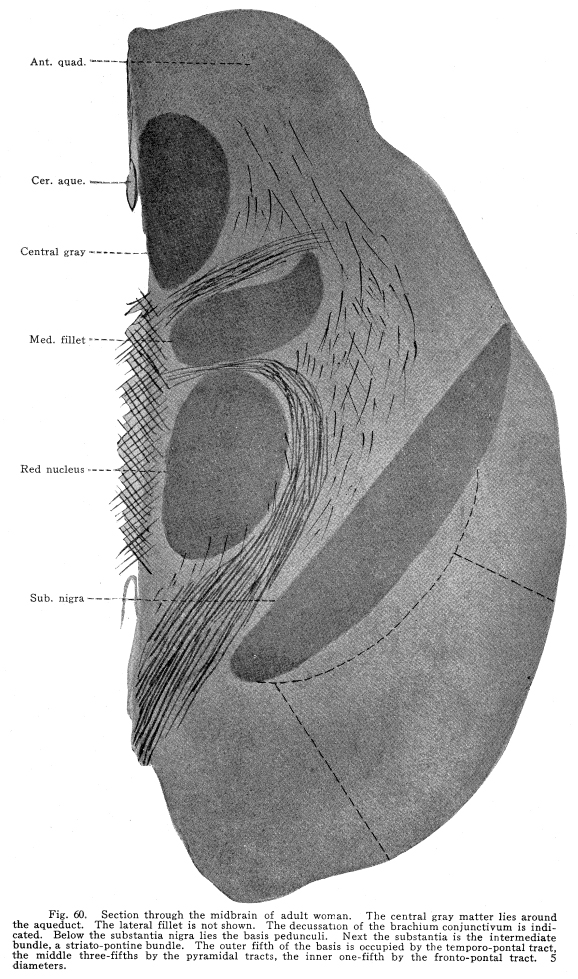
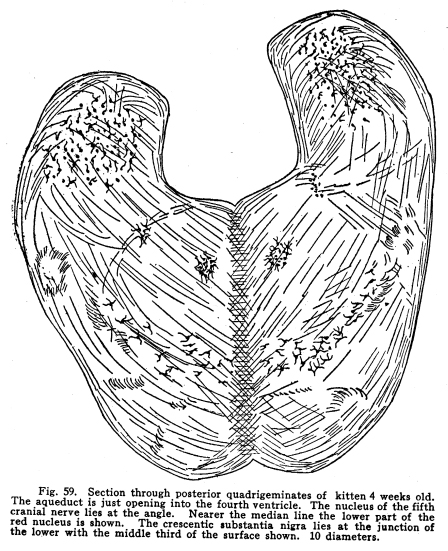
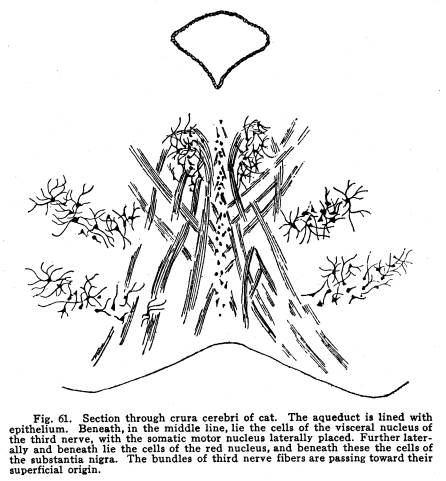
The inferior colliculus is composed of the two posterior quadrigeminate bodies. These bodies have a less complicated structure than do the superior bodies. (Fig. 59.) They are smaller, and contain only a single group of multipolar nerve cells. The brachium conjunctivum passes the inferior colliculus on its ventral and lateral aspects. The inferior colliculus receives a large part of the fibers from the lateral fillet. Thus the body is concerned in coordinating the reflexes of the auditory structures. It receives impulses from the following sources: I. The most important incoming stream of impulses is from the auditory nuclei by way of the lateral fillet. A few fibers, the axons of the cochlear ganglion cells, pass directly to the inferior colliculus. Other fibers are axons of the terminal nuclei of the auditory nerve, others are from the superior olive and the nucleus of the trapezoid body. II. Fibers from the internal geniculate body and thalamic nuclei pass to the posterior quadrigemina by way of the posterior brachium. These also are concerned in the coordination of the auditory reflexes. III. Descending fibers from the temporal lobes terminate in the inferior quadrigemina. These are concerned in carrying the efferent impulses to the auditory structures, as homologous fibers are concerned in the nutrition of the orbital tissues. Fig. 60. Section through the midbrain of adult woman. The central gray matter lies around the aqueduct. The lateral fillet is not shown. The decussation of the brachium conjunctivum is indicated. Below the substantia nigra lies the basis pedunculi. Next the substantia is the intermediate bundle, a striato-pontine bundle. The outer fifth of the basis is occupied by the temporo-pontal tract, the middle three-fifths by the pyramidal tracts, the inner one-fifth by the fronto-pontal tract. 5 diameters. Ast. Quad.; Cer. Aque.; Central gray; Med. Fillet; Red nucleus; Sub. Nigra; IV. Fibers from the medial fillet enter the posterior quadrigemina. These are probably reminiscent fibers, since the earlier colliculus received somatic sensory and viscero-sensory fibers. Fig. 59. Section through posterior quadrigeminates of kitten 4 weeks old. The aqueduct is just opening into the fourth ventricle. The nucleus of the fifth cranial nerve lies at the angle. Nearer the median line the lower part of the red nucleus is shown. The crescentic substantia nigra lies at the junction of the lower with the middle third of the surface shown. 10 diameters. V. Ascending fibers from the dentate nucleus of the cerebellum reach the body. These probably carry impulses of two classes: those of the indirect visceral sensory path, which must be largely reminiscent, and the impulses from the cerebellum itself, concerned in the coordination of the ear muscle movements with the movements of the muscles of other parts of the body. The posterior quadrigemina send impulses to the following structures: I. Impulses pass to the motor nuclei of the trigeminal and facial nerves, especially to those portions of these nuclei which control the ear muscles, but also to the portions which control other muscles of the head and face. II. Impulses pass to the nuclei of the oculo-motor, the trochlear and the abducens. In this way the movements of the ear and the eye are properly related to one another. III. Impulses pass to the vaso-motor center in the medulla. In this way the circulation through the ear tissues is in part controlled. IV. Impulses are carried by the tecto-spinal tract to the spinal cord at all levels. These fibers, while perhaps slightly functional in maintaining muscular tone and the equilibrium of the body, are probably chiefly reminiscent. V. Fibers from the inferior colliculus pass downward to the nuclei of the auditory nerve to the cochlea. In part the impulses carried by this path are relayed in the auditory nuclei. VI. Impulses from the inferior colliculus are carried to other related ganglia and nuclei. The quadrigeminate bodies are not often diseased. Their arterial supply and venous drainage are not easily impeded. They are supplied with blood by the ganglionic arteries, branches from the arterial circle of the cerebrum (circle of Willis). They are functional, and thus evade the tendency to disease characteristic of rudimentary structures. Tumors, gummata and tubercular nodules may affect the quadrigeminates, though rarely. The diagnosis is difficult during life, except in very limited lesions involving recognizable functional centers. Injuries affecting the superior colliculus are followed ultimately by degeneration of the retina, and, to a certain extent, of the other orbital tissues. There is immediately a lack of coordinate movements of the eyeballs. The manner in which the eyeball is displaced varies according to the location of the lesion and the manner in which the motor nuclei of the cranial nerves are also affected. The unequal dilatation of the pupils may help in diagnosis in some cases of unilateral lesions. Beneath the aqueduct the midbrain is composed of two fairly well-differentiated parts. The external layer is of white matter; it extends from the ventral surface to the crescent of pigmented gray matter called the substantia nigra. This external layer of white matter is called the basis pedunculi, or the crusta, in the older terminology. The bases pedunculi are of comparatively recent origin. In the lower vertebrates the tegmentum and the collicular region only are found. The process of cephalization is associated with the development of the longer fiber tracts which transmit impulses upward and downward. The base pedunculi are made up of these longer fiber tracts, which become necessary with the progressive development of the striate bodies and the cerebral hemispheres. The bases pedunculi are each composed of three main groups of fibers. These are not to be recognized as different in any way during adult normal life, but they are distinguished by their differing myelinization periods, and by the fact that they undergo degeneration after lesions of different areas of the cortex. (Fig. 60.) The outer one-fifth of each basis is composed of fibers called temporo-pontal. They are (1) the axons of cells in the temporal lobe of the cortex and the adjacent areas of the occipital lobe, or (2) axons of cells of the lenticular nucleus, with which the cortical axons form synapses. These axons terminate in the nucleus pontis; from the nucleus pontis the impulses are transmitted to the cerebellum. This is one of the indirect motor pathways. A few fibers pass directly to the cranial nerve motor nuclei. The median one-fifth of the basis pedunculi is composed of fibers from the frontal lobe to the pons. The axons of the cells of the frontal lobes pass directly by way of this tract to the pons, or they terminate in the gray matter of the thalamus or the striate body, from which axons pass by the same path to the pons. Thus the median and the lateral one-fifth of the basis is composed of fibers carrying impulses from the cortical centers to the pons, whence the impulses are referred to the cerebellum, and from there to the lower motor segments. The remaining three-fifths of the basis, placed in the middle of crescent, give place for the pyramidal fibers. These are the axons chiefly of the large pyramidal cells of the pre-central gyrus. These axons descend through the pons and medulla, giving off fibers to the red nucleus, substantia nigra, nuclei of the reticulum, and to the motor nuclei of the cranial nerves, to the lower part of the medulla. Here about four-fifths of the fibers decussate, and pass on the opposite side of the cord, to be distributed mostly to the centers of the somatic motor nerves of the lower part of the body and the legs. The remaining fibers pass on the same side of the cord until they reach the segment of their termination, when they also decussate, and are distributed chiefly to the centers for the control of the somatic muscles of the upper part of the body and the arms. Above the three divisions just mentioned lies a strand of fibers called the intermediate bundle. It is homologous with the external bundles of fibers. It is composed of the axons of cells of the striate bodies, and terminates, for the most part, in the nucleus pontis. Fig. 61. Section through crura cerebri of cat. The aqueduct is lined with epithelium. Beneath, in the middle line, lie the cells of the visceral nucleus of the third nerve, with the somatic motor nucleus laterally placed. Further laterally and beneath lie the cells of the red nucleus, and beneath these the cells of the substantia nigra. The bundles of third nerve fibers are passing toward their superficial origin. The
intermediate bundle is represented, to a certain extent, in the lower vertebrates.
It offers transmission to the few descending fibers from the developing
striate bodies, and probably is concerned in bringing the movements of
the body into relationship with the sensory impulses from the olfactory
centers.
The tegmentum is that portion of the midbrain included between the substantia nigra and the aqueduct. It is continuous with the tegmental region of the pons and medulla caudad, and with the tegmental region of the thalamus cephalad. The tegmentum is composed of fiber tracts, among which lie several groups of nerve cells. For
the most part the fiber tracts of the tegmentum are ascending. (Fig.
61.) The following have been studied:
The medial fillet (lemniscus) is composed of the axons of cells of the nucleus gracilis, nucleus cuneatus, and the nuclei of all the cranial sensory nerves except the cochlear portion of the auditory nerves. All of these fibers decussate, so that the fillet of one side carries sensory impulses from the opposite side of the body. The axons of the nucleus cuneatus and nucleus gracilis pass as internal arcuate fibers just dorsal to the inferior olive, decussate, and turn abruptly forward through the medulla, pons and midbrain. In passing they give some fibers to the motor nuclei of the cranial nerves, and a bundle of considerable size, the superior fillet, to the anterior quadrigemina. A few fibers stop at the inferior quadrigemina. The terminations in the quadrigemina are probably largely reminiscent, though a certain amount of movement of the eye and ear muscles in answer to the bodily sensations may thus be secured. The
fibers of the fillet terminate, for the most part, in the optic thalamus.
Probably the main termination for the medial fillet is the lateral thalamic
nucleus, though some investigators describe a different course for those
axons which arise from the cuneatus (globus pallidus, partly of the same,
partly of the opposite side).
The lateral fillet is the auditory pathway through the midbrain. It is composed of axons of the cells in the (1) cochlear nuclei, chiefly of the opposite side; (2) nucleus of the trapezoid body, chiefly of the same side; (3) superior olivary body of both sides; (4) nucleus of the lateral fillet of the same side; (5) inferior quadrigeminate body of the same side. These fibers pass together to the medial geniculate body, with whose cells the fibers form synapses. In passing the corpora quadrigemina the lateral fillet gives off fibers to these centersa large bundle to the posterior colliculi, by means of which the ear movements are in part coordinated and a few fibers to the superior collilculi, by means of which the movements of the eye and ear are in part related in function. The fibers of the brachium conjunctivum (superior cerebellar peduncle) are composed chiefly of the axons of the nucleus dentatus. These decussate for the most part just beneath the aqueduct, and pass forward to the red nucleus, where they terminate. A few pass onward to the lateral nucleus of the thalamus. The
brachium conjunctivum contains also a few fibers which are axons of the
cells of the red nucleus, which pass to the nucleus dentatus of the cerebellum.
This tract is that which apparently arises from the anterior ascending cerebello-spinal tract in the neighborhood of the the brachium conjunctivum, though in reality it is probably an individual tract throughout, and only intermingled with the ascending cerebello-spinal in passing upward through the cord. The fibers of this tract are axons of the cells of the posterior horns of the cord, or the dorsal nucleus, or both, with which the entering axons of the posterior roots form synapses. These axons decussate, or the axons of the posterior horn cells decussate, in the neighborhood of the segment of the entering root fibers. The tract is so closely intermingled with the anterior ascending cerebello-spinal tract that it is not possible to distinguish them except in degenerating material. The tract passes with the cerebello-spinal to the neighborhood of the brachium conjunctivum, where the cerebellar turns caudad to enter the cerebellum, while the spino-thalamic fibers continue cephalad to the thalamus. In passing through the medulla and pons a few fibers from the nuclei of the cranial nerves of common sensation join the tract and ascend with it. It gives off a few fibers to the superior and the inferior colliculi, which are probably reminiscent, and a few to the substantia nigra, whose function is not known. The chief termination of the tract is in the lateral nucleus of the thalamus. This tract carries impulses aroused on the opposite side of the body by temperature changes and painful stimulations, together with a part of the tactile impulses. The
tegmentum carries also a tract containing both ascending and descending
fibers, and a few descending tracts.
The medial longitudinal bundle (posterior longitudinal bundle) is composed of an ascending and a descending fasciculus. The ascending part is continuous and homologous with the anterior fasciculus proprius. It is composed of the axons of cells of the posterior and postero-lateral and central regions of the spinal gray matter, and in the terminal nuclei of the cranial sensory nerves, and probably in the nucleus gracilis and nucleus cuneatus. These fibers decussate, chiefly, and terminate about the cells of the motor nuclei of the cranial nerves, both somatic motor and visceral motor, in the quadrigeminate bodies, and a few fibers pass to the lateral nucleus of the thalamus. It is chiefly concerned in the transmission of those impulses necessary to the coordination of the complex reflex actions, including the muscles of the trunk, limbs and head. This tract also includes axons of cells in each of the nuclei of the third, fourth and sixth cranial nerves which pass to each of the others of the group, both of the same and of the opposite sides. The
descending strand of fibers is composed of axons of the cells of the superior
lateral nucleus of the reticular formation. In the pons it receives
the axons from the middle lateral nucleus, and the superior, middle and
inferior central nuclei of the reticular formation, and in the medulla
the axons of the inferior lateral nucleus of the reticular formation.
These fibers pass in the anterior fasciculus proprius of the cord throughout
its length (ponto-spinal tract), decreasing in size all the way as it gives
off fibers to the lateral part of the gray crescent at all levels.
In the medulla and pons and midbrain it gives fibers to the cranial motor
nuclei.
This
is almost exclusively a descending tract. Its fibers are the axons
of the cells of the quadrigemina, chiefly the anterior colliculus.
The fibers decussate almost completely, and pass caudad through the reticular
formation into the antero-medial fasciculus proprius of the cord.
It gives off most of its fibers to the nuclei of the third, fourth and
sixth cranial nerves and to the region of the cilio-spinal center in the
upper thoracic cord, but others of its fibers pass through the entire length
of the cord, giving off fibers in each segment. This bundle includes
very few fibers other than those of the tecto-spinal tract.
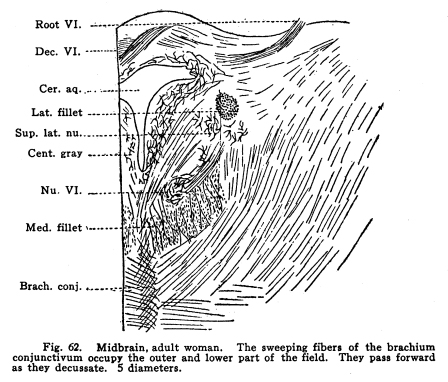
This is a tract whose function is not known. It is composed of axons of cells probably in the globus pallidus of the lenticular nucleus, and it terminates in the inferior olivary nucleus. Experimental stimulation of the globus pallidus gives no perceptible results, and no accounts of clinic evidence are on record. The rubro-spinal tract is composed of the axons of the cells of the red nucleus, which descend to the center of the gray crescent of all levels of the spinal cord. The tract probably carries with it axons of the cells of the subthalamic nuleus (Luys) and of the substantia nigra. These fibers decussate and pass downward through the tegmentum, pons, medulla and cord, giving off axons and collaterals to the motor nuclei, both somatic and visceral, and to the various centers scattered through the gray matter of these structures, including those concerned in both visceral and somatic coordinations. This tract, with the posterior longitudinal bundle, probably carry the impulses concerned in the emotional and instinctive reactions. The motor nucleus of the trigeminal nerve lies in the gray matter under the aqueduct. Its uppermost fibers arise near the opening of the third ventricle, but for the most part they come from the floor of the aqueduct near the opening of the fourth ventricle. These fibers are continuous with the motor fibers of the trigeminal through the pons. The gray matter of the midbrain includes all or a part of the nuclei of the third, fourth, fifth and sixth cranial nerves. These are discussed elsewhere. Fig. 62. Midbrain, adult woman. The sweeping fibers of the brachium conjunctivum occupy the outer and lower part of the field. They pass forward as they decussate. 5 diameters. Root VI.; Dec. VI.; Cer. Aq.; Lat. Fillet; Sup. Lat. Nu.; Cent. Gray; Nu. VI;Med. Fillet; Brach. Conj.
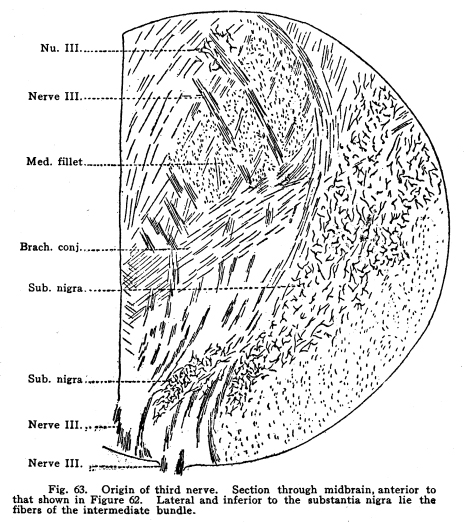
Fig. 63. Origin of third nerve.
Section through midbrain, anterior to that shown in Figure 62. Lateral
and inferior to the substantia nigra lie the fibers of the intermediate
bundle. Nu. III; Nerve III; Med. Fillet; Brach. Conj; Sub. Nigra; Sub.
Nigra; Nerve III; Nerve III.
This is a mass of gray matter, containing many cells, which are pigmented. It is a crescentic body, which forms the boundary between the tegmentum and the basis pedunculi. (Figs. 60, 62, 63.) The substantia nigra receives fibers, chiefly collaterals, apparently, from the following sources: I. From the medial fillet. II. From the lateral fillet. III. From the spino-thalamic tract. IV. From the brachium conjunctivum. V. From the red nucleus. VI. From the corpora striata by way of the intermediate tract. VII. From the pyramidal tracts. VIII. From the optic thalamus. IX. From the corpora mammillaria. The
impulses from the substantia nigra are carried, for the most part, by way
of the medial longitudinal bundle to the nuclei of the reticular formation.
Other connections are with the cranial nerve motor nuclei, the cerebellum
by way of the brachium conjunctivum, probably the olivary nucleus, and
there is also a direct path with the rubro-spinal tract to the spinal segments
through the length of the cord.
The
region between the red nucleus and the substantia nigra and the nucleus
hypothalamicus is occupied by a mass of interlacing fibers with cells interspersed
among them. These cells are rather large, multipolar, and have long,
freely-branching dendrites. The destination of their axons is not
known, but some of them seem to join the axons of the red nucleus cells
and those of the substantia nigra. The cells of the zona incerta
are to be considered, probably, as part of the centers for the coordination
of the movements of the emotional and instinctive reactions.
The nucleus hypothalamicus (Luys body) lies dorso-lateral to the substantia nigra, with which it appears to be continuous in the cat and the dog. This nucleus resembles the red nucleus in structure. Its cells are polymorphic and multipolar. Some Gogli cells of Type II are included. Into this nucleus pass fibers from the fillets, from the ascending tracts of the tegmentum to a certain extent, from the red nucleus and the substantia nigra. Descending fibers from the globus pallidus terminate in this nuleus. The axons of the intrinsic cells of the nucleus pass to the cerebral cortex, to the thalamus, to the red nucleus and substantia nigra, and to the nuclei of the reticular formation. Some investigators have described fibers from Luys body to the nucleus dentatus. Axons of the nucleus hypothalamicus enter the putamen and terminate by forming synapses with the cells of that center. This is not to be considered as a center. It is here mentioned only on account of its relationship with neighboring centers, and on account of its phylogenetic interest. It is a small body, about one-third of an inch in height and about half as great in its diameters. It takes its name from a fancied resemblance to a pine cone. It lies between and just anterior to the anterior quadrigeminates. It is formed as an outgrowth from the inferior portion of the superior walls of the third ventricle. It is composed of two lamina of gray matter, a number of solid cords of glandlike cells, blood vessels, and many small concretions called brain sand. Many fine, non-medullated nerve fibers, probably sympathetic, and derived from the superior cervical ganglion, branch freely among the nervous matter and the glandular tissue. The peduncles of the gland carry fibers, probably passing in both directions, to the nucleus habenulae, the anterior nucleus of the thalamus, and perhaps other nuclei. The pineal gland is rarely diseased; when it becomes subject to tumors (sarcomatous, fibroid, possibly adenoid) or an excessive accumulation of the brain sand, or becomes degenerated or injured through the disease of neighboring structures, no symptoms referable to the gland are present. Experimental stimulation of the gland, and the experimental injection of its extract into the blood of other animals, produces no results. This structure is reminiscent of the middle eye of certain lower forms. In certain reptiles and amphibia it is functional; in others rudimentary retina, lens and nerve fibers are demonstrated. |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}