
|
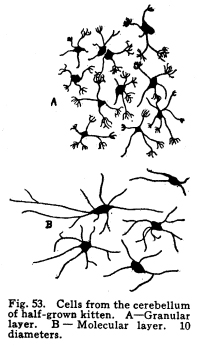
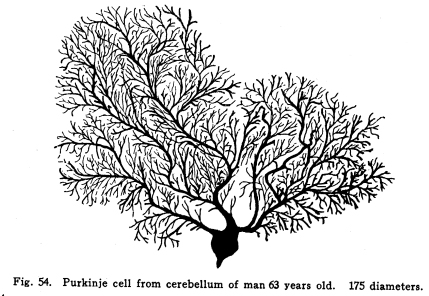
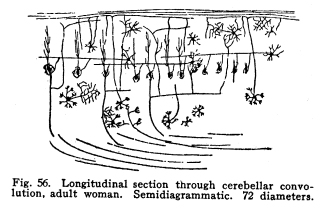
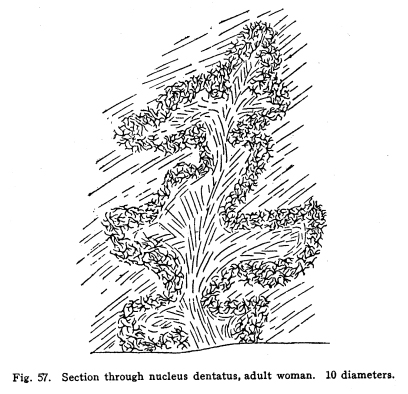
The cerebellum together with the pons represents the full development of the metencephalon. It is a fair representative of the complexity of relationship which characterizes those nerve centers whose functions have been subject to changes during their phylogenetic development. In the lowest vertebrates the metencephalon consists of a segment scarcely more differentiated than is any other segment of the neural axis. A prolongation from the lateral aspect of the metencephalon represents the beginning of a cerebellum. It receives, at first, only the terminals or collaterals from those sensory cells of the second order which receive the impulses from the lateral line sensory organs. (These organs, it should be remembered, are represented in the adult mammal only by certain of the structures of the middle ear.) In fishes of a somewhat higher order the cerebellum receives also terminals and collaterals from the secondary gustatory nucleus. Later, the fibers from the mesencephalic nuclei, concerned in the reception of impulses of sight and hearing, and from the gracile and cuneate nuclei, concerned in the reception of the sensory impulses from the body tissues, are received. The ascending cerebellar fasciculi are of more recent origin. Among fishes the cerebellum is very well developed, especially in the direction of the coordination of the sensory impulses. The life habits of fishes require this delicate coordination in order that they may retain their position while floating in a medium of about the same specific gravity as their own bodies. With the beginning of the land habits in amphibia the cerebellum becomes a less conspicuous object in the nervous system. The disappearance of the lateral line organs causes a large part of the nerve fibers to the cerebellum to be lost. With the more exact vision and the more need for reactions to answer speedily the stimulation received from a single sense organ, the need for wide coordination of sensory impulses becomes superseded by the need for a more exact coordination of efferent impulses. Thus it happens that we have the appearance of an organ of great efficiency in one class of vertebrates, becoming of less importance in another, and later reaching an efficiency along slightly different lines, far surpassing the primitive development. From the lower amphibians to man the development of the cerebellum has progressed in the direction of increasing the coordination of the efferent rather than the afferent impulses. This end is secured, in part, by means of the very wide sensory connections, the foundation for which was laid in the fish cerebellum, in part by the development of more distant sensory connections, and in part by the development of other connections with higher centers, and by the increasing complexity of the relations of all of these. The structure of the cells of the cerebellum itself has changed comparatively little. The axons of the Purkinje cells remain the sole efferent path of impulses from the cerebellar cortex. The incoming impulses, whether from sensory nuclei, lower centers, midbrain, or the cerebral cortex itself, are, so far as our present knowledge can say, limited to the primitive simple methods of receiving; the incoming fibers either terminate in brushlike endings in connection with the brushlike endings of the granule dendrites, or they exhaust themselves by fine branchings, which apply themselves to the dendrites of the Purkinje cells. It is not possible to localize the fibers from the different parts of the nervous system in the cerebellum with any exactness. Fig. 53. Cells from the cerebellum of half-grown kitten. A Granular layers. B Molecular layer. 10 diameters. Fig. 54. Purkinje cell from cerebellum of man 63 years old. 175 diameters. Fig. 55. Section across cerebellar convolution, adult woman. Semidiagrammatic. 72 diameters. Fig. 56. Longitudinal section through cerebellar convolution, adult woman. Semidiagrammatic. 72 diameters. The histological structure of the cerebellum is as complex as its phylogenetic development would indicate. The cortex presents two layers, between which lie the bodies of the Purkinje cells. These cells are sometimes classified as a third, or ganglionic layer. The outer or molecular layer is so called from the presence of a number of very small nerve cells. (Fig. 53.) The cells of the outermost part of this layer are very small multipolar cells, whose axons exhaust themselves by repeated branchings in the immediate neighborhood of the cell body. They are a form of the Golgi Type II cells. In the deeper part of the molecular layer are found the stellate or basket cells. These are of stellate form, multipolar, with freely branching dendrites. Their axons pass along the deeper part of the molecular layer, in a direction parallel with the direction of the folds of the cerebellar cortex. Thus, in order to secure sections showing these axons it is necessary to cut the convolutions of the cerebellum longitudinally. (Fig. 56) From these axons collaterals are given off, which descend to the bodies of the Purkinje cells and break up into a feltwork of fibrillae, which make a basket around the bottle-shaped bodies of the Purkinjes. (Fig. 18.) The number of collaterals from any given axon of the basket cell seems to be very large. (I have seen seven collaterals from one such axon in one section of Golgi material.) The molecular layer contains a very large number of nerve fibers. These include the following: Axons from the small granules of the granular layer pass through the molecular layer to the periphery, divide in a T-shaped manner, and pass tangentially along the cortex. From these branching axons collaterals descend into the molecular layer to form synapses with the nerve cells therein. The dendrites of the Purkinje cells branch freely among the cells of the molecular layer. The axons of the Purkinje cells give off collaterals, which pass toward the cerebellar cortex through the molecular layer, and form synapses with the multipolar cells. In the molecular layer are also the climbing fibers of the cerebellum. The middle or ganglionic layer, as it is sometimes called, is composed of the bodies of the Purkinje cells. (Fig. 54.) These are large and bottle-shaped, and their bodies are closely invested by the basketlike branchings of the collaterals of the axons of the stellate or basket cells of the molecular layer. The dendrites of the Purkinje cells are characteristic. They branch very freely in one plane without the molecular layer. The dendrites are beautifully tree-shaped, and the whole cell presents a very striking appearance in sections prepared after the method of Golgi. The plane of division of the dendrites is placed at right angles to the direction of the convolutions, so that if one wishes sections showing the dendrites of the Purkinje cells, it is necessary to cut the convolutions crosswise. (Fig. 55.) The dendrites of the Purkinje cells are closely followed by the branchings of the climbing fibers. The inner granular layer is so called from the appearance of its most abundant cells. These are called granules, and include two classes, large and small granules. The granule cells are alike in form. They are multipolar, and their dendrites terminate in peculiar, brushlike end tufts. (Fig. 53.) These end tufts are in close relationship with the similar brushlike terminations of the collaterals and terminals of the moss fibers of the cerebellum. The small granule cells have long axons, which penetrate the molecular layer, branch in a T-shaped manner, and run tangentially along the cortex, giving off collaterals to the cells of the molecular layer at intervals. The large granules are of the Golgi Type II class. Their axons exhaust themselves by repeated branchings among the dendrites and axons of their immediate neighborhood. Through the granular layer the axons of the Purkinje cells pass as they leave the cortex; through this layer also the collaterals from these axons pass toward the cortex. The entering climbing and moss fibers penetrate this layer in reaching their destination. Beneath the granular layer is found the cerebellar medulla of white fibers passing to and from the cortex. These fibers are supported by neuroglia cells. This white matter includes fibers of two classes, neither of which is as well known as is desirable. The climbing fibers enter the cerebellum, but their origin is not certainly known. They probably enter by way of the restiform body or the branchium pontis. They pass to the cortex, branch freely, and apply themselves to the dendrites of the Purkinje cells, which they follow to their ultimate extremities. The moss fibers enter the cerebellum, but their origin is not known. They branch freely, and their collaterals and probably also their terminals break up into the brushlike end tufts already described for the granule cell dendrites, and closely applied to them. The axons of the Purkinje cells pass downward through the white matter, and they alone are corticifugal. It is not certainly known whether most of the Purkinje cells terminate in the dentate nucleus, or whether most of them leave the cerebellum. Part of them certainly stop in the dentate nucleus. When it is remembered that the dendrites of the Purkinje cells branch widely in a plane at right angles to the direction of the convolutions, that these widely-branching dendrites are thoroughly permeated with the collaterals and axons of the multipolar cells of the molecular layer, with the tangential fibers of the axons of the small granular cells, and with the axons of Golgi Type II cells; when it is remembered also that the axons of the stellate cells pass parallel with the direction of the convolutions, and thus at right angles to the direction of the Purkinje dendritic plane, and that a considerable number of Purkinje cells may receive the basketlike terminations of any one stellate axon, then the complexity of this structure may in part be realized. The recurrent collaterals of the Purkinje axons add to the intricacy of the arrangement. Fig. 57. Section through nucleus dentatus, adult woman. 10 diameters. Fig. 58. Cells of nucleus dentatus from same section as Figure 57. 800 diameters. The masses of gray matter centrally placed require much more study. The nucleus dentatus is the largest of these masses. It is found in the lower animals as a single mass of gray matter of simple form. In the human cerebellum it presents the characteristic dentate appearance, very much like that of the olive. Its phylogenetic development is not known. It may represent an infolding of the cerebellar cortex. (Figs. 57, 58.) The nucleus emboliformis is placed at the hilum of the dentatus, like a cork for that structure. The globosus lies mesially from the emboliformis. These nuclei present some evidence of being of phylogenetic descent from the secondary gustatory nucleus of fishes. The fibers of the anterior ascending cerebellar fasciculus pass into these nuclei, but whether the axons terminate in part or as a tract, or whether they pass without relay to the cortex, is not certainly known. The nucleus tectis lies in the roof of the V-shaped ventricle. It seems to represent an infolding of the cortex. The cerebellum receives nerve impulses from very nearly all parts of the central nervous system, and since this includes all or practically all of the sensory neurons, both visceral and somatic, it follows that the cerebellum receives impulses from the entire body. The connections of the cerebellum are as follows: I. Somatic sensory neurons of the first order may send axons or collaterals to the superior vermis by way of the restiform body. This includes the fibers of the fasciculus gracilia and fasciculus cuneatus, which pass into the restiform body without relay, and the fibers of cranial sensory nerves, which may follow the same path. II. Visceral sensory neurons of the first order may send axons or collaterals to the cerebellum by way of the restiform body. Probably these include very few fibers. III. Somatic sensory neurons of the second order send many axons and collaterals to the cerebellum, chiefly by way of the restiform body from the nucleus gracilis, nucleus cuneatus and the cranial nerve nuclei of termination. These seem to reach the cortex of the superior vermis. IV. Visceral sensory neurons of the second order send many axons to the superior vermis cortex. These include the fibers of the anterior ascending cerebellar fasciculus (direct cerebellar), which passes by the restiform body, and a part of the posterior ascending cerebellar fasciculus (Gowers tract), which passes through the medulla and pons and send some fibers by the brachium conjunctivum (superior peduncle) into the cerebellum. V. Axons or collaterals from the nucleus pontis pass to the cortex of the hemispheres of the cerebellum. For the most part these axons pass to the contra-lateral hemisphere, but a few of them pass to the hemisphere of the same side. VI. Axons of cells of the red nucleus have been said to enter the cerebellum by way of the brachium conjunctivum. This connection has been doubted by later studies. It seems probably that a small number of fibers form the red nucleus to the cerebellum pass by way of a bundle in the brachium conjunctivum. VII. The cells of the olive send axons to the cerebellum, but whether to the dentate nucleus or to the cortex is not known. The cerebellum sends impulses by almost as varied pathways. I. The axons from the cells of the nucleus dentatus make up the larger part, if not all, of the brachium conjunctivum (superior peduncle). These axons terminate for the most part, in the red nucleus, but also in part in the quadrigeminates and the optic thalamus. The brachium conjunctivum fibers decussate immediately after emerging from the cerebellum. These fibers probably carry impulses concerned in consciousness of heat, cold, pain, muscular effort and the various visceral sensations. II. Axons from the cells of the cortex of the cerebellar hemispheres pass by way of the brachium pontis (middle peduncle) to the nucleus pontis, mostly contra-lateral, but partly to the same side. III. Descending axons either from the cortex (the Purkinje cells) or from the dentate nucleus pass downward through the cord, forming synapses with the cells of the central and postero-lateral gray matter, and through them affecting the activity of the cells of the anterior and the lateral horns of the cord. IV.
Axons from either the cortex or the dentate nucleus pass by way of the
restiform bodies to the olive, the medullary centers, and the motor cranial
nerve nuclei. Very little is known of the various relationships.
The functions of the cerebellum have been the subject of much study. Experimental evidence is almost as much at variance with clinic evidence as the different manifestations of clinic evidence vary among themselves. It seems to be well proved that the cerebellum is chiefly concerned in the maintenance of the tome of the visceral and somatic muscles, the maintenance of the tome of the visceral and somatic muscles, the maintenance of equilibrium, the coordination of complex movements, and with the transmission of visceral and certain common somatic sensations to the cortex. These functions are of great importance, yet widespread disease of the cerebellum may be associated with few or no localizing symptoms. In the presence of tumors or other lesions involving the cerebellar hemispheres it is rare for a diagnosis to be made ante-mortem unless the peduncles are involved. Lesions of the vermis are sometimes recognized, but it sometimes happens that even vermis lesions escape recognition until postmortem. Stimulation of the cerebellar hemispheres in the anesthetized animal produces no perceptible effects. Stimulation of the central gray matter may be followed by contraction of the muscles, chiefly those of extension of the same side of the body, if the basal ganglia remain intact. Section of the brachium conjunctivum precludes this reaction. Stimulation of the brachium pontis is followed by contraction of the muscles, chiefly those of extension of the same side of the body. These movements include the diaphragm and the intercostals. No visceral effects have been seen to follow any stimulation of the cerebellar tissues. Stimulation of the restiform body produces no perceptible effects. Diseases of the cerebellum are rather rare and are not always to be recognized. The gross lesions include those found in any other part of the brainabscesses, gummata, tubercles, etc., and the symptoms produced are often not very well related to the normal functions of the cerebellum, as indicated by its structural relationships. The removal or injury of a considerable part of the cerebellum is followed by a loss of muscular power and of coordination on the same side of the body. There is no perceptible loss of sensation. There may be or may not be muscular tremors and athetosis. If the injury is not progressive, and the subject is not too old, there is later produced some effort at compensation, either upon the part of the cerebellum left intact, or by other centers capable of coordinating the impulses concerned in producing the complex actions of the skeletal muscles. No symptoms referable to an injury of the viscero-sensory paths have been described in cerebellar disease. The cerebellum, as one of the nerve centers, must depend for its normal activity upon the same conditions which affect the other nerve centers; that is, the cerebellum depends for its food supply upon the blood brought to it. This blood must be good, clean, and kept rapidly flowing if the cerebellar cells are to maintain their proper activity. This activity also depends upon the receipt of certain impulses from the other parts of the nervous system already described. In the lack of these impulses, or if they should be rendered abnormal in any way, the normal coordinations are not found. It is doubtless true that many of the mistakes made by weary people, or by people whose bodies are in any way subnormal, may be due to one of two conditionseither the cerebellar cells are poorly nourished and are thus inefficient, or the nerves carrying sensory impulses, or other impulses, to the cerebellum are not such as to lay the foundation for normal coordinating activities in the cerebellar cells. It is by means of this relation that many of the accidents called careless are produced by persons who are neurotic or poorly nourished, or whose nerve cells have a lower liminal value than usual for any reason. During childhood the progressive development of the neurons concerned in the higher unconscious coordinations may be associated with periods of poor coordinations. Children at this age are, no doubt, often blamed for the awkwardness and carelessness, due, in large part, to the delayed or unbalanced development of the cerebellar neurons, or of the neurons upon whose activity the coordinations depend. |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}