
|
The medulla is a mass of nerve tissue which lies caudad to the pons and cephalad to the upper extremity of the spinal cord. It is about an inch in length and about half an inch in its antero-posterior diameter. At its lower extremity it has the transverse diameter of the spinal cord, with which it is directly continuous; this is about half an inch. At its upper extremity it has a transverse diameter of nearly an inch, as has the pons, with which it also is continuous. Its anterior aspect presents a series of grooves and ridges, which pass from above downward. The posterior surface of the medulla forms the floor of the lower triangle of the fourth ventricle. Its anterior and lateral surfaces are broken by the roots of the cranial nerves. The
white matter of the medulla is composed of ascending, descending, transverse
and antero-posterior fibers. A few tracts contain both ascending
and descending fibers.
The ascending fibers are mostly continuous with those of the cord; the descending fibers, for the most part, are continuations of the same tracts in the pons. The ascending tracts are as follows: I. The medial fillet is composed of the axons of the cells of the nucleus gracilis and the nucleus cuneatus. These axons pass as internal arcuate fibers anteriorly, and pass behind the olivary body. They decussate, turn forward, and pass through the substance of the medulla on their way to the optic thalamus. As they traverse the medulla and pons the axons of terminal nuclei of the sensory cranial nerves (except the auditory) are added to the fillet, and with it pass forward to the optic thalamus. This tract carries the finer sensations of touch and muscular effort. II. The ascending anterior cerebello-spinal tract is continuous with the same tract in the cord. It passes through the medulla and pons to the region of the brachiuim conjunctivum, where it turns backward to enter the cerebellum. It is composed of the axons of the cells of the dorsal nucleus of the opposite side of the cord, and during its pathway through the medulla it receives axons of the terminal nuclei of the cranial nerves of the opposite side. It transmits impulses initiated by temperature changes, painful stimuli, sensations of muscular effort, and the visceral sensations. A part of the tactile impulses are carried by this tract. III. The spino-thalamic tract is so closely associated with the tract just mentioned that it is not possible to separate them in the pons. That they are physiologically separate is certain. The fibers of the spino-thalamic tract arise from the cells of the dorsal nucleus of the opposite side, pass with the anterior ascending cerebello-spinal tract to the angle of that tract; the spino-thalamic tract then proceeds on its way to the optic thalamus. It carries the impulses of pain and temperature, but probably not of muscular effort. A part of the tactile sensations are probably carried by this tract. IV. In the lower part of the medulla the fasciculus gracilis and the fasciculus cuneatus are found. They are the axons of the cells of the sensory ganglia, and they terminate, for the most part, in the nucleus gracilis and the nucleus cuneatus. A few fibers pass directly into the restiform body. V.
In the lower part of the medulla the posterior ascending cerebello-spinal
tract is found. It is composed of the axons of cells in the nucleus
dorsalis, mostly of the same side, and it enters the restiform body on
its way to the superior vermis. It carries visceral sensory impulses,
and perhaps also impulses from other structures.
I. The pyramids occupy a prominent position in sections of the medulla. They are placed very near the anterior surface of the medulla. Near the lower extremity of the medulla about four-fifths of the fibers decussate just under the surface. The decussation is easily seen from the anterior aspect of the medulla. The other one-fifth of the pyramidal fibers remain upon the same side until they approach the segment of their termination in the cord. The fibers of the pyramidal tracts are the axons of large pyramidal cells of the precentral convolutions of the cerebral cortex of the same side (before the decussation). They are distributed to the central gray matter of the crescent of the opposite side of the cord, and to the motor nuclei of the cranial nerves of the opposite side. II. The anterior longitudinal bundle (tecto-spinal tract) is continuous with the same tract in the pons and in the cord. It is composed of axons of the cells of the colliculus. It terminates in the center of the crescent of the spinal cord. Many of its fibers terminate in the cilio-spinal center in the upper part of the thoracic cord. It carries the impulses concerned in the reflex actions initiated by visual impulses, and perhaps other sensory impulses. III. The rubro-spinal tract is continuous with the same tract in the pons and in the spinal cord. It is composed of the axons of the cells of the red nucleus and probably of neighboring gray matter, and it terminates in the center of the crescent of the spinal cord, and in the nuclei of the cranial motor nerves. IV. The spinal tract of the trigeminal nerve is composed of the descending limb of the T-shaped division of the entering trigeminal sensory roots. The axons are in the semi-lunar ganglion (Gasserian ganglion). The fibers terminate in the substantia gelatinous of the posterior horn of the cord of the upper cervical segments, and in the nucleus of termination of the fifth nerve in the medulla. V.
The tractus solitarius is a bundle of fibers composed of descending fibers
from the sensory part of the seventh, ninth and tenth cranial nerves.
It passes to the centers of the lower medulla and upper cervical cord,
and is supposed to be concerned in carrying the impulses of the respiratory
reflexes.
I. The medial (posterior) longitudinal bundle is continuous with the anterior fasciculus proprius of the cord. Through this tract the axons of cells in lower nuclei pass to enter into higher nuclei, and axons of cells in higher nuclei pass toward cells of the lower nuclei. Thus these fibers are associational in function. Part of the ascending fibers of this tract in the cord are joined by other fibers from the sensory nuclei of the cranial nerves, and of these some terminate in other nuclei of the sensory and motor cranial nerves, while others pass to the optic thalamus. The descending part of the medial longitudinal bundle is composed of the ponto-spinal tracts. These are axons of cells of the reticulum of the pons, and they terminate by forming synapses with the cells of cranial motor nerves and the anterior horns of the cord. Part of the ponto-spinal fibers are crossed, part are not crossed. II. The lateral fasciculus proprius is made up of ascending and descending fibers, which bring the different levels of the medulla into functional relationship. Among these fibers lies the descending olivary bundle, which is supposed to carry impulses from the centers of the pons and the basal ganglia to the olive, and the olive-spinal tract, which is supposed to carry impulses from the olive to the spinal centers. III. The restiform body contains a number of ascending and descendidng fibers. These are: (a)
The cerebello-spinal ascending and descending fibers; (b) the external
arcuate fibers, which pass from the nucleus gracilis and nucleus cuneatus
of the opposite side, chiefly; (c) fibers from the inferior olive to the
cerebellum and from the cerebellum to the inferior olive.
The transverse and antero-posterior fibers may be grouped for convenience in description. I. The axons of the nucleus gracilis and nucleus cuneatus pass as anterior and posterior external arcuate fibers toward the anterior part of the medulla. The posterior set surround the olive of the same side, pass to the opposite side, and enter the restiform body. The anterior set pass between the olives, surround the olive of the opposite side, and enter the restiform body. Both sets of external accuate fibers give off some fibers to the olives and the arcuate nuclei both of the same and the opposite sides. The decussation of the fillet causes these fibers to appear as horizontal fibers. II. In their decessation the pyramidal fibers pass transversely for a short distance. III. The medullary striae are axons of the cells of the auditory nuclei. They pass from these nuclei, placed far laterally as they are, toward the median raphe. Here they plunge into the substance of the medulla and form synapses with the cells of the various secondary auditory nuclei. IV. The root bundles of the vestibular, hypoglossal, glosso-pharayngeal, vagus and spinal accessory nerves are seen as antero-posterior fibers. V.
Various small bundles of horizontal fibers connect the nuclei of each level
in functional relationship. This connection varies according to the
physiological requirements of the different nuclei.
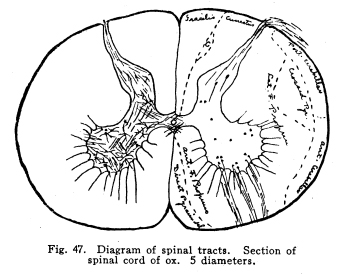
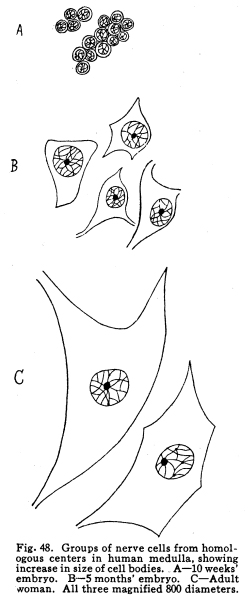
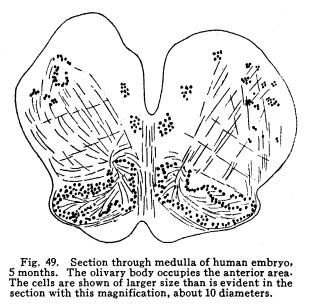
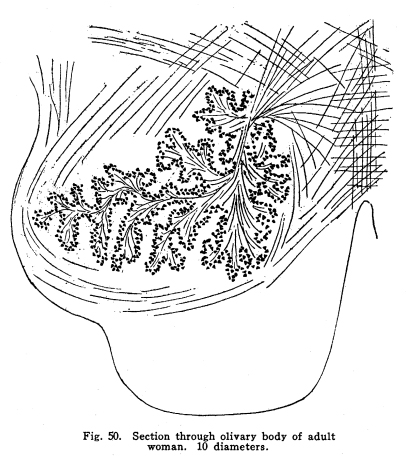
The centers for the trigeminal, facial, acustic, glosso-pharyngeal, vagus, accessory and hypoglossal nerves are found in part or completely in the medulla. These centers are discussed elsewhere, and will not be considered at this time. This group of nerve cells lies upon the anterior aspect of the medulla. It is continuous with the nucleus pontis, with which it is probably homologous. Some of the external arcuate fibers form synapses with the cells of this nucleus, and it seems probable that other fibers from the medullary nuclei may also terminate in this nucleus. This relation is not certainly known, but in sections a number of fibers apparently from the nucleus of the trigeminal and from unidentified cell groups of the medulla seem to enter the arcuate nuclei. (Fig. 48.) This structure is found as such in mammals only. Masses of nerve cells of similar arrangement and position are found in lower vertebrates,s even among fishes. By some authors these centers are held to be homologous with the olive of the higher vertebrates. (Figs. 49, 50.) The statement frequently made that the olivary nucleus does not appear until the sixth month of embryonic life does not seem true. (In a fetus of five months supposed existence I have found the olivary nucleus fairly well developed.) Fig. 47. Diagram of spinal tracts. Section of spinal cord of os. 5 diameters. Fig. 48. Groups of nerve cells from homologous centers in human medulla, showing increase in size of cell bodies. A 10 weeks embr;yo. B 5 months embryo. C Adult woman. All three magnified 800 diameters. Fig. 49. Section through medulla of human embryo, 5 months. The olivary body occupies the anterior area. The cells are shown of larger size than is evident in the section with this magnification, about 30 diameters. Fig. 50. Section through olivary body of adult woman. 10 diameters. Two accessory olivary bodies are found. The olive has functions similar to those of the cerebellum, with which it is very closely related in structure. Lesions of the olive are associated with loss of coordination, ataxia of the cerebellar type, usually nausea, and other symptoms of cerebellar disease. Antemortem diagnosis of olivary disease is probably impossible, in the absence of symptoms indicative of localization of surrounding structures. Fibers enter the olivary nucleus from the following sources: I. Axons of the nucleus gracilis and the nucleus cuneatus enter the olivary nuclei of both sides. Probably other secondary sensory axons also enter the body. II. Faibers from the cerebellar hemispheres pass by way of the restiform body to the contra-lateral olive. III. Descending fibers, probably from the lentiform nucleus, terminate in the olive. Fibers
leaving the olivary nucleus pass to the cerebellum, either to the nucleus
dentatus or to the hemispheres, chiefly of the opposite side. Other
fibers from the olivary nucleus pass downward through the cord to enter
into gray matter of the various levels through its whole extent.
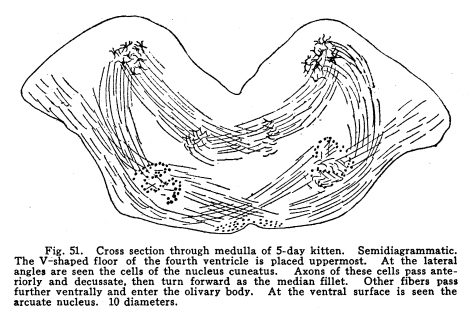
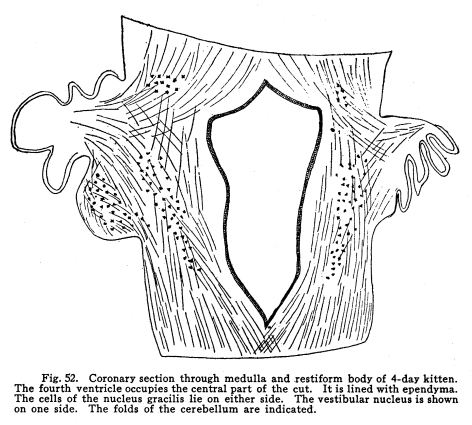
These nuclei may be considered as one. Indeed, the gray matter of which they are composed is continuous. They differ only in the fact that the cuneatus receives the sensory impulses form the upper part of the body, while the gracilis receives the impulses from the lower part of the body. Their central connections are similar. Fig. 51. Cross section through medulla of 50day kitten. Semidiagrammatic. The V-shaped floor of the fourth ventricle is placed uppermost. At the lateral angles are seen the cells of the nucleus cuneatus. Axons of these cells pass anteriorly and decussate, then turn forward as the median fillet. Other fibers pass farther ventrally and enter the olivary body. At the ventral surface is seen the arcuate nucleus. 10 diameters. Fig. 52. Coronary section through medulla and restiform body of 4-day kitten. The fourth ventricle occupies the central part of the cat. It is lined with ependyma. The cells of the nucleus gracilis lie on either side. The vestibular nucleus is shown on one side. The folds of the cerebellum are indicated. These nuclei receive the ascending fibers which are the axons of cells of the sensory ganglion on the posterior roots of the cord, and of cells in the posterior horns of the cord with which the entering axons form synapses. The impulses thus transmitted are concerned in the sensations of touch and muscular effort, especially such of these sensations as are concerned in delicate perceptions, in exact localizations, and which are speedily recognized. The axons of these nuclei have the following destinations: I. As internal arcuate fibers the axons pass through the medulla, decussate, and turn forward as the medial fillet. This is the most direct path of the sensory impulses to the centers concerned in consciousness. The fillet terminates in the optic thalamus, after giving fibers to the cranial nerve nuclei and the colliculi. II. Axons pass as external arcuate fibers, both anterior and posterior, around the olives and into the cerebellum by way of the restiform body. From these tracts fibers are given off to the olives and to the arcuate nuclei. (Fig. 51.) III. Axons pass into the restiform body and the cerebellum directly. (Fig. 52.) IV.
Axons pass anteriorly to the nuclei of the cranial nerves.
This nucleus is homologous with the nuclei of the pontine reticulum. It receives fibers from the anterior ascending cerebellar tract, the nucleus gracilis and the nucleus cuneatus, and probably also from the terminal nuclei of the cranial sensory nerves and from the corpora mammillaria. Its axons pass into the cerebrum by way of the brachium conjunctivum, and to the spinal cord with the ponto-spinal fibers in the anterior longitudinal bundle. The pons is a structure of irregularly cylindrical outline. It is about an inch long, a little more than an inch broad, and about an inch in its antero-posterior diameter. Laterally,
the transverse fibers which appear upon the surface of the pons gather
together and form the brachium pontis, or middle cerebellar peduncle.
Its anterior aspect presents only a mass of transverse fibers. Its
posterior aspect forms the floor of the upper triangle of the fourth ventricle.
Its superior boundary is continuous with the midbrain, its inferior with
the medulla.
The pons is composed of a very complex network of fibers and groups of nerve cells. The bundles of fibers are variously separated and combined, so that it is not easy to trace even the well-recognized tracts through the rete mirable. The fibers which enter into the formation of this complex network are as follows: I. The pyramidal tracts descend through the anterior portion of the pons. These fibers are the axons of the neurons of the central gyri, and are on their way to the spinal and medullary centers. The decussation of these fibers occurs in the medulla, it will be remembered, so that injuries to the pyramidal tracts in this location produce the symptoms peculiar to lesions of the cortex or capsule, so far as the character of the paralysis is concerned. II. The tecto-spinal fibers pass through the pons on their way from the quadrigeminate bodies to the spinal and medullary centers. III. The rubro-spinal fibers pass through the pons on their way from the red nucleus, and probably also from the substantia nigra and the sub-thalamic region, to the spinal and medullary centers. All of these descending tracts send terminals and collaterals to the pontine centers. By means of this connection the coordination of the lower centers is maintained, in part, and also the action of the sixth and fifth nerve nuclei are controlled. IV. Fibers from the red nucleus and other structures related to it in function send many fibers to the nucleus pontis, and thus enter into relationship with the cerebellum. V.
Fibers from the temporal, occipital and frontal lobes of the cerebrum send
fibers to the pontine nuclei, and thus enter into relationship with both
the cerebellum and the lower spinal medullary centers.
I. The median fillet passes near the posterior surface of the pons, near its lateral aspect. It is composed of the axons from cells in the nucleus gracilis, nucleus cuneatus, and the terminal nuclei of the sensory cranial nerves as they pass to the optic thalamus. It will be remembered that this tract gives off fibers also to the quadrigeminates and perhaps to other neighboring nuclei. II. The lateral filler lies lateral and posterior to the median fillet. It is composed of axons of the cells of the auditory nuclei, and also a few fibers directly from the vestibular and cochlear ganglia. The cells of the nucleus of the trapezoid body send fibers to the lateral fillet. The fibers of the trapezoid body pass into the lateral fillet also. The lateral fillet passes upward through the pons to the optic thalamus, and sends many fibers to the posterior colliculus. III.
The anterior ascending cerebello-spinal fasciculus passes through the pons
on its way to the cerebellum. This tract arises from the cells of
the dorsal nucleus (Clarkes column) in the cord, runs through the neural
axis to the neighborhood of the brachium conjunctivum, turns backward,
and enters the cerebellum to terminate in the cortex of the superior vermis.
As this tract passes through the pons it gives off a bundle of fibers with
which it has been very closely intermingled in its entire previous course.
This bundle, the spino-thalamic tract, is not to be distinguished from
the anterior ascending cerebello-spinal tract until their division in the
region of the brachium conjunctivum; but the fact that it is physiologically
a distinct tract is shown by the effects of lesions involving the two division;
the spino-thalamic tract evidently carries impulses of pain and temperature
exclusively, and the tactile sense, in part; the anterior ascending cerebello-spinal
carries impulses of muscular sense, chiefly.
I. The posterior or medial longitudinal bundle lies near the posterior part of the medial raphe. It includes both ascending and descending fibers, which carry impulses from the various nuclei of the cranial nerves and their related nuclei to other cranial nerve nuclei. II. The anterior longitudinal bundle is physiologically homologous with that just mentioned. It carries the fibers which transmit the impulses from the different centers concerned in the movements of the eyeball to other centers having the same or related functions. The tecto-spinal tract is carried with it. III.
Unidentified fiber groups may be concerned in transmitting impulses upward
and downward to and from the various nuclei and centers in the pons, and
both above and below the pons.
I. The ventral portion of the pons is occupied by large masses of fibers, which pass transversely across and through the structure. These are of two classes. The axons of cells in the cerebellum, chiefly the Purkinje cells of the lateral hemispheres, but probably partly the cells of the dentate nucleus, form synapses with the cells of the pontine nuclei, chiefly of the opposite side, but partly of the same side. Axons of the cells of the pontine nuclei pass to the cerebellar hemispheres, chiefly of the opposite side, but partly of the same side. II. The fibers of the corpus trapezoideum are a conspicuous feature of pontine sections. The fibers of this body arise form the different auditory nuclei and pass to the opposite side of the pons. They turn abruptly forward and form the lateral fillet. About midway from the raphe to the angle the nucleus of the trapezoid body is interposed, and in this body, and in the superior olive and its accessory nuclei, the fibers are interrupted. The axons of the cells of these bodies assist in the formation of the lateral fillet. III.
A number of fibers pass from the nuclei of either side of the pons to form
synapses with symmetrical or related nuclei of the opposite side.
The antero-posterior bundles of the pons include those fibers which associate the different nuclei, those collaterals and terminals from the ascending and descending tracts which enter or leave the nuclei placed anteriorly or posteriorly to them, and the efferent fibers of the motor nerves as they seek their places of emergence from the surfaces. The term nucleus pontis is used to include the masses of cells scattered through the ventral part of the pons. It is considered as a single nucleus, on each side, whose cells have been separated by the transverse fibers of the pons and by the descending bundles of the pyramidal, fronto-pontal, temporo-pontal and intermediate tracts. The cells of this nucleus are large multipolar cells, surrounded by the interlacing fibrillae of the incoming axons, which thus form synapses with them. These incoming fibers include the axons of cells in the frontal and temporal lobes of the cerebral cortex, and probably from other cortical areas, and from the basal ganglia. The pyramidal tracts give off collaterals and terminals to the nucleus pontis also. The
axons of the nucleus pontis pass for the most part to the contra-lateral
cerebellar hemisphere. A few pass to the cerebellar hemisphere of
the same side.
In
the reticular formation of the pons are found small scattered groups of
cells. These have been called, from their position, lateral median,
superior central, median central, and inferior central. The cells
of these nuclei are large and multipolar, and their bodies and dendrites
are surrounded by the branching fibrillae of the incoming fibers.
The axons of these cells bifurcate into an ascending and a descending branch.
The destination of the ascending branches is not known. The descending
branches pass downward through the pons, medulla and spinal cord.
They terminate at different levels in these structures by forming synapses
with the cells of the motor nerve nuclei to the lowermost spinal centers.
These ponto-spinal tracts undergo partial decussation in the medulla.
The nucleus of the trapezoid body includes a number of scattered cell groups within the trapezoid fibers. The fibers of the trapezoid body and medullary striae, axons of the auditory nuclei, form synapses with the small multipolar cells of this nucleus. The axons of the cells of the nucleus pass with the uninterrupted fibers from the auditory nuclei to form the lateral fillet. The axons from the auditory nuclei terminate by forming a cuplike expansion upon the bodies of the cells of the trapezoid nucleus. This form of synapsis is not described for other centers. The
sensory nuclei of the trigeminal and auditory nerves are found in part
within the pons. The motor part of the trigeminal and the facial,
and the nuclei of the trochlear, abducens and a part of the oculo-motor
nerves, are also found within the pons. The description of these
centers is given in connection with the discussion of the relations of
the cranial nerves.
The
olivary group of nuclei includes the superior olive, the semilunar nucleus,
and the preolivary nucleus. These cell groups contain small multipolar
nerve cells. They receive axons from the auditory nuclei of both
sides and send axons into the lateral fillet. They send axons also
to the nuclei of the abducens, trochlear and oculo-motor nerves.
Thus the nuclei form a part of the auditory-ocular reflex arc. The
movements of the eyeballs in answer to sudden sounds are in part mediated
by way of this pathway.
The functions of the pons are thus seen to be largely in the nature of correlation. The pons forms part of the pathway to and from the cord, cerebrum and cerebellum. It is chiefly functional in the transmission, but certain of the coordinations and the reflexes are mediated within its gray matter. Diseases of the pons are rare, and are not apt to be recognized antemortem. The injury of any perceptible area of the pons is associated with immediate death; the injury of the cranial nerve nuclei is associated with the symptoms referable to the loss of function of these nerves. The injury to the pontine nucleus may be associated with incoordination of movements, or, if the lesion be gradually produced, no localizing symptoms may be found. Injuries of the ascending or descending tracts produce the symptoms due to the loss of the corresponding functions. |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}