
|
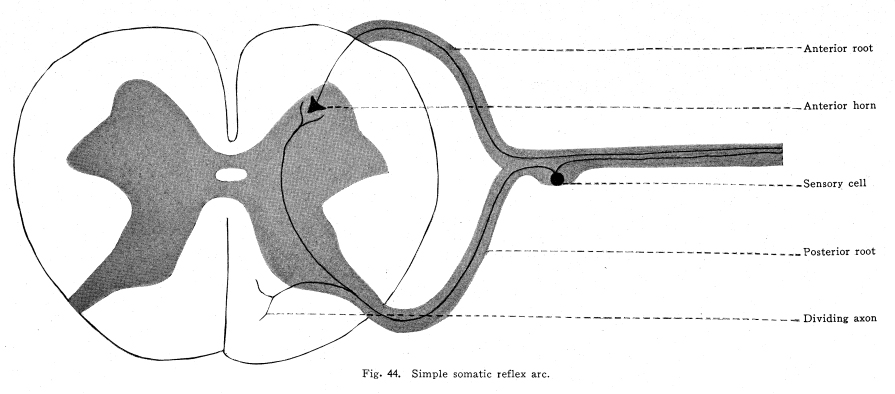
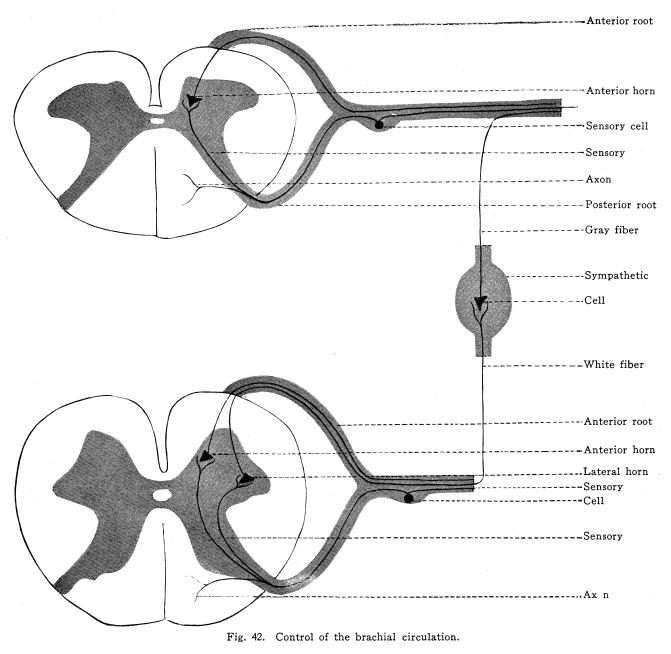
The spinal cord is to be considered in regard to two chief functions. From the standpoint of the lower vertebrates, the spinal cord is chiefly a series of neuron groups, which are centers for the control of all of the skeletal and visceral structures of the corresponding segment. Later development shows a progressive development of long tracts. These long tracts serve several purposes, all concerned in increasing the efficiency of the central nervous system in its duties in unifying the parts of the body. The long tracts may be afferent, in which case they supply the higher centers with the nerve impulses needful in their functions; or they may be efferent, in which case they carry to the spinal centers the impulses sent out from the higher centers in answer to the action of the impulses transmitted by the afferent tracts; or they may be associational, in which case they enable the different centers, both in the cord and in certain of the higher centers, to act in unity. The spinal centers have retained in great degree their original functions in controlling the structures of the corresponding segments of the body, through all the great development of the long tracts. The spinal centers are not to be considered, with their present development, at least, to be separate and distinct from an anatomical standpoint. It is not possible to select from the cells of the cord the exact groups concerned in any reaction. It is not possible, at present, to say certainly whether the same nerve cell or group of cells may be concerned in the coordination of the nerve impulses of more than one different function. There are no facts of interest in this connection except the symptoms produced by certain degenerative processes in the cord, and the results of these observations are, so far, very contradictory. It is not possible to determine the exact extent of any nerve center. For if any center should extend through several segments of the cord, or if it should be very limited in extent, but should receive axons or collaterals from the sensory roots of several segments, the relations would be identical from a functional standpoint. From the symptoms observed in cases of disease following injury to the cells in the gray matter of the cord, it appears that the viscero-motor centers extend through several segments of the cord. Yet it is possible that they merely receive association fibers from several adjacent segments. It is possible, then, only to locate these centers rather vaguelythat is to say, that any one is found between a certain spinal segment and another lower spinal segment. The spinal cord includes that part of the central nervous system which lies within the vertebral canal. Its uppermost extremity is marked by the lower limit of the decussation of the pyramids, and by the uppermost fibers of the origin of the first cervical nerve. It terminates at the lower border of the first lumbar vertebra. The entire length of the cord is about seventeen or eighteen inches, according to the varying heights of individuals. Below the limit of the cord lies an extension continuous with it, the filum terminale. This resembles the cord for a short distance; it contains fibers and gray matter resembling embryonic nerve tissues. Vestiges of the coccygeal nerve roots are sometimes to be found. The filum terminale reaches down into the sacral region, where it terminates by becoming continuous with the connective tissue cord formed from the rudimentary meninges, and is attached to the sacrum. The spinal cord varies in diameter in different regions; it is a little more than a half inch in its largest diameter, and a little less than a half inch in its smallest diameter. Its weight is about one and one-half ounces, avoirdupois. In form the spinal cord is an irregular cylinder. It presents two swellings. The upper of these is called the cervical enlargement, which corresponds to the area of origin of the nerve roots which supply the arms and shoulder girdle. The lower is the lumbar enlargement, which gives rise to those nerves which supply the pelvic girdle and the legs. The spinal cord contains at its approximate center a canal called the sixth ventricle, or the neural canal, or the central canal of the cord. It is lined with epithelium, which is ciliated in the embryo. The gray matter of the cord surrounds this central canal. It is arranged in the form of a very irregular H, whose arms vary in length and breadth in the different areas of the cord. Throughout the thoracic region, and to slight extent in certain other regions, there is found a slight swelling upon the central part of each lateral arm of the H-shaped gray matter. This is called, when present, the lateral horn of the cord. (Figs. 35, 36, 37.) That part of the gray matter which extends forward is called the anterior horn, that which extends backward is called the posterior horn. The anterior horn includes two classes of cells. The large multipolar cells, which are so conspicuous a feature in cord sections, are the motor cells of the first order; their axons pass by way of the anterior roots to the skeletal muscles and carry the nerve impulses which cause the movements of the skeleton. (Fig. 44.) Associated closely with these cells are others, smaller and also multipolar, whose axons branch very freely close to the cell, and do not leave the gray matter. These cells (Golgi Type II) are functional in assisting in the coordination and transmission of impulses concerned in reflex actions. The cells of the lateral horn send axons by way of the anterior roots and the white rami communicantes to the sympathetic ganglia. They carry impulses which are concerned in the control of the viscera and the blood vessels. (Figs. 39, 40, 41, 45.) The posterior horn includes cells of several functions. In this part of the gray matter are found cells whose axons terminate by branching among the cells of the other parts of the gray matter. These are associational in function. Others of the posterior cells send axons upward and downward through the gray matter or through the tracts of white matter at the edge of the gray, to upper or lower segments. They terminate by entering the cord and forming synapses with different cells at that level. The nucleus dorsalis (Clarkes column) is found only in the thoracic cord. It sends axons by way of the anterior ascending cerebello-spinal tract and by way of the posterior ascending cerebello-spinal tract to the cortex of the superior vermis of the cerebellum, or to its central nuclei. This tract carries the impulses concerned in muscular sense. Axons from this column of cells probably make up the spino-thalamic tract, which carries impulses concerned in pain and the temperature sense to the optic thalamus. The white matter of the cord is composed of ascending, descending, and mixed tracts. The descending tracts are as follows: I. The direct pyramidal tract extends from the medulla to the mid-dorsal region. It terminates by forming synapses with the cells in the central region of the opposite spinal crescent. It is composed of axons of the large pyramidal cells of the precentral convolutions of the same side of the cerebral cortex, and it carries impulses concerned in the voluntary movements of the arms and shoulder muscles. Fig. 42. Control of the brachial circulation. Anterior root; Anterior horn; Sensory cell; Sensory; Axon; Posterior root; Gray fiber; Sympathetic; Cell; White fiber; Anterior root; Anterior horn; Lateral horn; Sensory; Cell; Sensory; Axon. II. The crossed pyramidal tract extends through the entire length of the cord. It terminates by forming synapses with the cells of the gray matter of the central part of the crescent of the same side. The tract is composed of the axons of the large pyramidal cells of the precentral convolution of the opposite side of the cerebral cortex, and it carries the impulses concerned in the voluntary movements of the lower part of the body. III. The rubro-spinal tract extends through the length of the cord. It is composed of fibers which are axons of the cells of the red nucleus, and probably the substantia nigra and the sub-thalamic region, and it terminates by forming synapses with the cells of the lateral horns and the central gray matter. Perhaps the fibers may pass in part to the anterior horns directly, but this is not certainly known. This tract carries impulses concerned in the emotional and instinctive movements of the skeletal muscles, and in the visceral activities associated with emotional states. IV. The anterior longitudinal bundle (tecto-spinal tract) extends through the length of the cord. Its fibers are axons of cells in the colliculi, and it terminates with the pyramidal fibers. It is supposed to be concerned in maintenance of the muscular tone, and certainly transmits impulses functional in the coordination of those reflex movements associated with sights and sounds. V. The olivo-spinal tract is not well known. Its fibers are the axons of cells in the inferior olive, and the impulses are supposed to be concerned in the coordination of the more complex movements of the body. It is closely related to the cerebellum in function. VI. The ponto-spinal tracts extend the length of the cord. They are composed of the axons of cells in the reticulum of the pons, which are distributed to the gray matter with the pyramidal tracts. The impulses carried by these tracts are probably concerned in the coordination of the muscular movements and in the maintenance of the tone of the skeletal muscles. The lateral ponto-spinal tract is decussated, the medial tract is not decussated. VII. The vestibulo-spinal tract has its origin in the vetibular nuclei. It is rudimentary structure in the higher vertebrates, but represents a connection of great importance in fishes. It forms synapses either with the anterior horn cells, or with cells closely related to these. The impulses carried by this tract are concerned in the maintenance of muscular tone, and the coordination of the movements associated with equilibrium. VIII. The descending cerebello-spinal tract extends through the length of the cord. Its fibers terminate by forming synapses with the cells of different regions of the gray matter of the cord at different levels. Its fibers are axons from the cerebellum. It is not known whether they are axons of the Purkinje cells or of the cells of the dentate nucleus. The impulses carried by this tract are concerned in maintaining the tone of both skeletal and visceral muscles, and in securing the coordinate activity of the skeletal muscles, especially those concerned in equilibrium. It is not known whether these impulses are greatly concerned in the visceral activities or not. The phylogenetic history of the relations seems to indicate the possibility that this function may be included. IX. The comma tract, though a descending tract, is derived from the sensory neurons. The entering axons of the sensory neurons divide into two branches, the shorter of which passes downward in the posterior funiculi for a distance of one or two spinal segments, and terminates by forming synapses with the cells of the lower levels. The impulses thus transmitted are probably concerned in the more complex reflexes, and in securing more perfect coordination of the movements of the skeletal and visceral muscles. The ascending tracts of the cord include the following: I. The fasciculus gracilis (tract or column of Goll) lies near the posterior median septum. It is composed chiefly of the axons of the sensory ganglion cells, and a smaller number of axons from the posterior horn cells from below the mid-dorsal region. The fibers pass to the nucleus gracilis in the medulla, where they terminate by forming synapses with the cells of that nucleus. A few fibers of this tract pass directly to the cerebellum by way of the restiform body. II. The fasciculus cuneatus (tract or column of Burdach) is composed chiefly of the axons of the cells of the sensory ganglia, and partly of a smaller number of axons of cells of the posterior horns. These fibers pass upward to the nucleus cuneatus in the medulla, and form synapses with the cells of that nucleus. A few of the fibers of this tract pass directly into the restiform body and into the cerebellum. III. There is some reason to consider the existence of a long sensory tract situated in the posterior gray commissure of the cord. This has been called Ciaglinskis tract. Its relations are not known. IV. The anterior ascendidng cerebello-spinal tract (Gowers tract) is composed of axons of the cells of the dorsal nucleus, chiefly of the opposite side. With it are carried the fibers of the spino-thalamic tract, which have the same origin. These tracts pass together into the region of the brachium conjunctivum, where the cerebellar part of the tract turns backward to enter the cerebellum, while the spino-thalamic tract passes onward to the optic thalamus. V. The posterior ascending cerebello-spinal tract (direct cerebellar) is composed of axons of the cells of the dorsal nucleus, chiefly of the same side. These fibers pass by way of the restiform body to the cerebellum, where they terminate either in the cortex of the superior vermis, or in the central gray matter of the cerebellum, by which the impulses are transmitted to the cortex of the superior vermis. VI. The median longitudinal bundle is composed of axons of the cells of the anterior horns, and it terminates in the cranial nerve nuclei and the thalamus. It is concerned in the more complex reflex actions. I. The antero-lateral fasciculus proprius lies next to the anterior horn on its lateral aspect. Its fibers are from the cells of the gray matter, and they pass both upward and downward, thus connecting the different levels of the same side of the cord. II. The antero-median fasciculus proprius lies next to the anterior horn on its central aspect. Its fibers pass both upward and downward to a certain extent, but chiefly they decussate in this passing, so that these fibers are concerned in connecting the contra-lateral segments of gray matter, both of the same and of neighboring levels. Within this tract lies the median longitudinal bundle. III. The posterior fasciculus proprius is made of axons which connect the different levels of the cord. They are mostly short fibers, only a few segments in length. The cornu-commissural tract is the postero-lateral bundle of fibers, which include also fibers concerned in connecting the contra-lateral gray matter, as well as the different homolateral levels of the cord. The centers of the upper cervical cord lie within the gray matter of the first, second, third and fourth cervical segments. The gray matter of this portion of the cord presents certain peculiarities. These segments occupy the upper part of the cervical enlargement. The posterior horn is capped by the substantia gelatinosa, among whose cells the descending branches of the fifth cranial nerve terminate. The ophthalmic division of the fifth supplies this region especially freely. The nucleus dorsalis (Clarkes column) is wanting. The intermedio-lateral cell group (lateral horn) is displaced anteriorly, probably because of the development of somatic functions and relations on the part of the cells whose axons make up the spinal part of the eleventh cranial nerve, and of those whose axons make up the phrenic nerve. The trapezius and the sterno-mastoid are innervated by the eleventh cranial, and the diaphragm by the phrenic. These muscles are especially subject to abnormal reflex contractions as the result of excessive viscero-sensory impulses from viscera, which are affected by pathological conditions. The anterior horn is large and broad. It includes several groups of cells. The mesial cell column is homologous with a similar group of cells through the whole extent of the cord, and it innervates the trunk muscles; in the case of the upper cervical group, the superior and inferior oblique, the rectus capitis posticus major and minor, the complexus and trachelo-mastoid, the splenius and semispinalis, the multifidus spinae, the transversalis cervicalis, cervicalis ascendens, by the posterior primary divisions, the splenius, platysma, scalene, longus colli, rectus capitis anticus major and minor, rectus capitus lateralis, and the sterno-mastoid in part by the anterior primary divisions. The hyoid muscles also are innervated from these segments, probably from a small cell group in the antero-lateral column. These cell groups are intricately associated with the cell groups of the posterior horns, and with the gray matter of neighboring segments. The muscles named above are thus affected by many of the sensory impulses reaching the segments. They are found abnormally contracted in many disorders of viscera, limb structures, and cranial structures. The antero-lateral cell group is not represented in the thoracic cord, but is well marked in the upper and lower cervicals; the axons of these cells innervate the limb muscles. These cell groups seem to represent a later phylogenetic development than the antero-mesial group; they are larger cells, have long axons, and are poorly associated with the other cell groups of the gray matter. This is, no doubt, the reason why the visceral reflexes have so little effect upon the limb muscles, though they are not altogether free from visceral reflexes, as is commonly supposed. They are very well supplied with association fibers from the sensory neurons innervating the limb muscles, skin and joint surfaces. The muscles innervated by the anterior lateral group are, Levator scapulae, teres major and minor, supraspinatus, and rhomboid. The somatic sensory cells of this group lie in the ganglia on the posterior roots of the corresponding nerves. The dendrites of these cells are distributed as follows: Skin over the back of the neck and the head to the vertex, including the mastoid process and part of the ear; the skin of the neck as far upward as the lower line of the mandible; the muscles innervated by the motor neurons of the same segments; the articular surfaces of the first, second, third, fourth and fifth vertebrae; and the sterno-clavicular and the acromio-clavicular articulations; the meninges of the corresponding spinal segments. No viscero-sensory neurons are found in these segments, but the viscero-sensory neurons of the fifth, eighth, ninth and tenth cranial nerves send collaterals and terminals to these segments. The sensory portion of the seventh is not known to send fibers to the cervical cord. The efferent impulses from these centers are influenced by both inhibiting and stimulating impulses from the following sources: I. The impulses from the skin, muscles and joint surfaces innervated by the somatic sensory neurons of the same segments affect the efferent impulses from these centers. By this relation the direct reflexes are governed. Under slightly abnormal conditions, such as slight malpositions of the articular surfaces mentioned, the contraction of the muscles may become excessive and constant, and these reflex muscular contractions may in turn send in sensory impulses which increase or decrease the abnormal position of the structural tissues, and the stream of sensory impulses may be further modified. II. Impulses from the sensory nuclei of the fifth, eighth, ninth and tenth cranial nerves are carried to the upper cervical centers. By this relationship abnormal conditions in the area of distribution of these nerves may cause excessive contractions of the muscles innervated by the upper cervical segments. Thus are produced the tension of these muscles during toothache, or gastritis, or pharyngitis, or otitis media, or conjunctivitis, etc. III. Impulses from the nucleus gracilis and nucleus cuneatus reach the upper cervical segments. At a result of this relationship abnormal sensory impulses from diseased viscera may cause abnormal tension of the upper cervical muscles. The tension of the upper cervical muscles so often found associated with diseased pelvic or abdominal organs is thus produced. IV. Impulses from the cerebellum, the vestibular nuclei, the quadrigeminates, and probably the nuclei of the pontine reticulum, reach the nuclei of the upper cervical segments. By this means the movements of the upper cervical muscles are coordinated, and their normal tone is maintained. V. The fibers of the direct pyramidal tract form synapses with the cells of the lateral cell mass of the opposite side, and these in turn send axons to the large motor cells of the anterior horn. Thus the impulses from the precentral cerebral convolution are enabled to carry the impulses concerned in the volitional control of the upper cervical muscles. VI. The rubro-spinal tract carries impulses from the red nucleus and the neighboring basal ganglia to these segments. By this means the upper cervical muscles are brought under the control of the emotional impulses. VII.
Descending impulses from the viscero-motor centers in the medulla reach
the upper cervical segments. By this means the movements of the diaphragm,
scaleni, etc., in respiration, and in the modified respiratory movements,
as coughing, sneezing, the respiratory factors in speech, etc., are controlled;
the diaphragmatic movements in vomiting, defecation, and similar actions
are secured.
This group of centers includes those situated in the fifth, sixth, seventh and eighth cervical segments, and the first thoracic segment of the cord. The gray matter in these segments presents certain peculiarities. The segments named include the lower part of the cervical enlargement. The posterior horn is capped by the substantia gelatinosa. In this region it is less conspicuous than in the upper cervical group, and it contains fewer nerve cells. There is a difference among neurologists concerning the place of the descending fibers of the fifth; by some authors it is given as reaching as low as the sixth cervical segment, while others consider the evidence insufficient to show fifth-nerve fibers below the second cervical segment. Reflexes initiated by the stimulation of the fifth nerve affect the action of the centers of the cord as low as the upper thoracic segment, but this may be due to the existence of closely-related association neurons. The lower cervical group evidently receives impulses from the fifth, whether by direct fiber paths or by means of interpolated neurons is not yet apparent. The nucleus dorsalis (Clarkes column) is found only in the seventh cervical and first thoracic segments. The viscero-motor column is scarcely to be found. A few fibers enter the phrenic from the upper part of the group, and a few assist in forming the spinal portion of the eleventh cranial nerve. Otherwise the column is not represented. The significance of these nerves is discussed in connection with the upper cervical group. The first thoracic and perhaps the seventh cervical segments contain a few cells of the cilio-spinal center. The anterior horns of this region are broad and large. The cells are arranged in groups, part of which are placed mesially and a part of which are placed laterally. The mesial group is homologous with a similar column of cells through the extent of the cord. Its fibers innervate the muscles of the trunk, including the semispinalis and multifidus spinae, the trachelomastoid, scaleni, longus colli, cervicalis ascendans, transversalis cervicis, complexus, and splenius. The cells in this group represent the older structure, phylogenetically, than the lateral group of cells; their central relations are complex, and they receive impulses from many sources, viscero-sensory, somatic sensory, and descending. The muscles named as being innervated from these cells are, therefore, subject to abnormal contractions as the result of excessive stimulation of the centers from other structures. It is especially noted that these contractions follow visceral disturbances of the cervical and cranial regions, as well as in the upper thorax. The lateral group of cells includes several groups, whose exact functions have not been certainly worked out. This column is not found in the thoracic region of the cord. Its cells send axons which innervate the muscles of the shoulder girdle and the arms, including the following: Teres major and minor, supraspinatus and infraspinatus, rhomboid, anconeus, subscapularis, serratus magnus, pectoralis major and minor, coracobrachialis, deltoid, biceps, triceps, brachialis anticus, supinators longus and brevis, latissimus dorsi, the pronators, extensors and flexors of the wrists and fingers, the lumbricales and interossei, and the thenar and palmar muscles. These cell groups represent the newer structure, phylogenetically; their central connections are not very freely associated with other cell groups, and the muscles named are not especially subject to abnormal contractions from sensory stimulations in other parts of the body. The somatic sensory cells lie in the sensory ganglia on the corresponding posterior roots. Their dendrites are distributed to the following structures: Skin over the arms, hands and fingers, and a small area over the anterior and posterior aspect of the thorax, innervated by the first thoracic nerve. The muscles innervated by the motor neurons of the same segments. The articular surfaces of the fifth, sixth and seventh cervical and the first thoracic vertebrae, the clavicle, the first costo-sternal, the scapula, the humerus, radius and ulna, and all the articulations of the wrist, hands and fingers; The meninges of the corresponding spinal segments. No viscero-sensory neurons are found in these segments, and the only sources of viscero-sensory impulses is by way of the descending fibers of the cranial nerves, and the association neurons of the thoracic segments. The efferent impulses from these centers are affected by both stimulatory and inhibitory impulses from the following sources: I. The somatic sensory impulses from the skin, articular surfaces, and muscles innervated from these segments affect the efferent impulses through simple and complex reflex action. Under slightly abnormal conditions, such as malpositions of the articular surfaces, or skin lesions, the contractions of the muscles may become constant and excessive. These muscular contractions may also affect efferent impulses. Since the structures innervated from the brachial plexus are phylogenetically of comparatively recent origin, the central associations are comparatively less complex and the reflexes are less severe and less frequent than in the case of certain other structures. II. Impulses from the cerebellum, the vestibular nucleus, the olive, the quadrigeminates, and probably the nuclei of the pontine reticulum, reach the lower cervical segments by way of the tracts named after their origin. By these connections the normal tone of the muscles innervated from the brachial plexus are maintained, and their movements are coordinated. III. Fibers from the direct pyramidal tract of the cord form synapses with the cells of the opposite side of the cord. By this means the voluntary impulses from the precentral convolutions reach the muscles of the opposite upper limb. IV. The rubro-spinal tract carries impulses from the red nucleus and adjacent ganglia to these segments. Thus the arm movements are controlled by the emotional reactions. V. Descending impulses from the viscero-motor centers in the medulla, and perhaps the pons and midbrain, reach these segments. The most conspicuous function thus mediated is that of respiration. |
{kind=link}
{kind=link}