
|
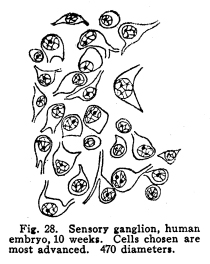
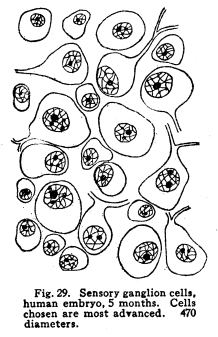
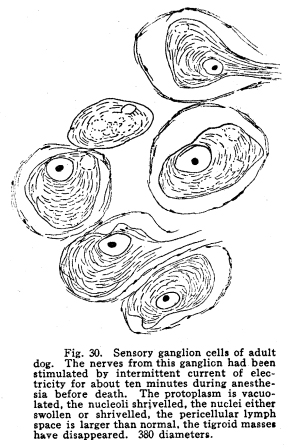
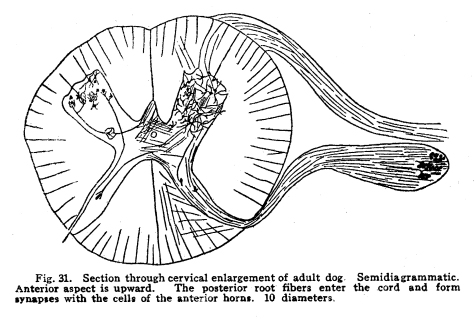
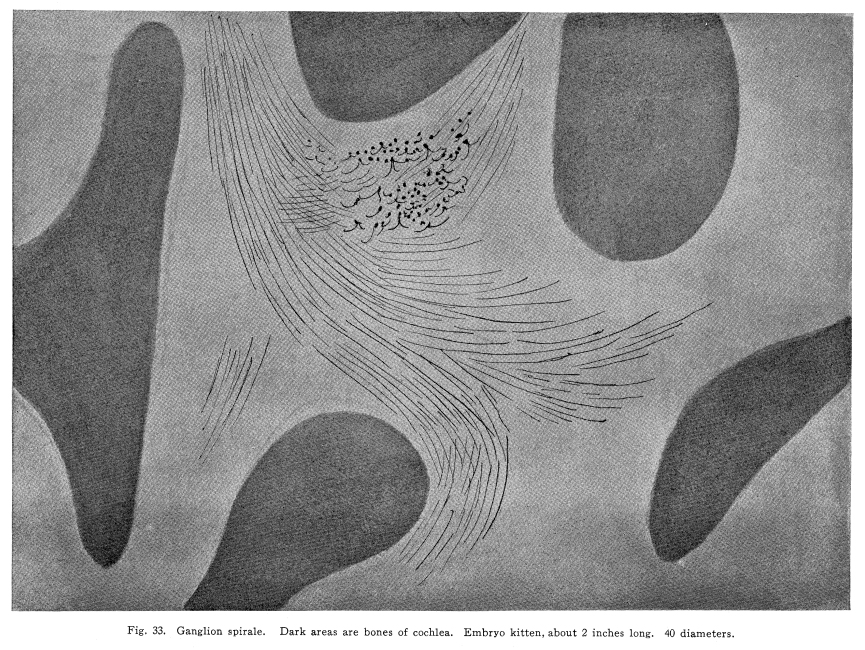
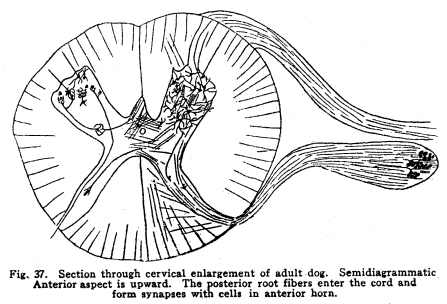
In order that the body may react to its environment in a manner which preserves the life of itself and its race, it is necessary that provision be made whereby the changes in the body itself and its environment may, either directly or indirectly, affect motor structures. In the case of the human being this relationship must include the consciousness of at least a part of the environmental and bodily conditions. This need for a functional relationship between the motor structures and those parts of the body most easily affected by external changes is met by the sensory neurons, the motor neurons, and various associational neurons by means of the which the various impulses from different parts of the body and from different sense organs are coordinated. The essential features of this function are: first, some form of peripheral structure capable of being affected by some character of environmental or bodily change; second, a nerve fiber and nerve cell capable of transforming the stimulus thus received into a nerve impulse; third, central relationships capable of transmitting these nerve impulses to the motor neurons; fourth, efferent neurons; fifth, motor structures capable of being affected by the nerve impulses transmitted to them. (Figs. 24, 25, 26, 27.) Sensory impulses form the foundation of all nervous activity. Practically every reaction to external or internal changes depends upon the transmission of sensory impulses aroused by these changes into the central nervous system. Mentality and volition are dependent upon sensory stimulation; the effects of any given sensory impulse may be immediate or may be long retained within the nervous system. But ultimately practically all efferent impulses are derived from precedent afferent impulses. Fig. 27. Sensory ganglion, adult dog. 200 diameters. Sensory impulses may initiate impulses to the skeletal or the visceral muscles or glands. These may pass over the simplest path, by way of the collaterals to the anterior or lateral horn cells, or they may follow more complex paths involving almost any number of neurons. The relation which the outgoing impulses bear to the incoming impulses depends upon the structural relationships of the neurons concerned, and upon the comparative liminal values of these neurons. Stimulation of the somatic sensory nerves of any segment of the cord may affect the activity of the skeletal muscles, the visceral muscles, the blood vessels, and the glands innervated from the same segment, either directly or by way of the sympathetic ganglia. Stimulation of the visceral sensory nerves of any segment of the cord may affect the same structures, according to the structural relationships and the comparative liminal values of the associational and motor neurons. The physiological value of these reactions is apparent. By means of the reflexes thus dependent upon the sensory impulses the repair of wounds is facilitated, rest of any injured part of the body is compelled, vaso-motor and viscero-motor impulses are governed in accordance with the needs of the body under slightly abnormal conditions, and adaptation and compensation are facilitated under those conditions in which recovery is impossible. These same reactions may, however, become a menace in the presence of long-continued sensory irritation, as in the habitual use of abnormal foods, slight malpositions of structural tissues, or any of the various peripheral irritations, such as eye strain, scar tissue in certain parts of the body, etc. The sensory impulses which are carried to the higher centers have no less importance in the bodily economy. The viscero-motor centers in the medulla depend in great part upon the sensory impulses carried by way of the long and the short spinal tracts, by the vagus and other sensory cranial nerves, and by the descending impulses from higher centers themselves ultimately dependent upon sensory impulses. Coordination is secured through the action of the cerebellar cells, which depend in like manner upon the sensory impulses carried by the ascending cerebellar tracts, the impulses from the cranial sensory nerves, and the descending impulses from higher centers which also are ultimately dependent upon the sensory impulses. Sensory neurons of the first order are those which are immediately affected by changes in the environment of the body, or by changes in the condition of the body tissues. It is not now possible to state how the environmental changes are able so to affect the sensory nerve endings as to initiate a nerve impulse. Since so great variations exist between the specific sensory nerve endings, it appears probable that the key to this problem may be found in a study of the structures and metabolism of these structures. A number of the varieties have been illustrated, but no study has yet been made of their metabolism, apart from the metabolism of the neuron. Very little attempt has been made to classify the nerve endings from a functional standpoint. There are thirty-one pairs of spinal sensory ganglia, and two pairs of rudimentary coccygeal ganglia. These ganglia are placed upon the posterior roots of the spinal nerves, and are situated within the corresponding intervertebral foramina. They are supplied by blood vessels from the corresponding veins and arteries, and the caliber of these vessels is controlled by vaso-motor nerves from sympathetic ganglia of the same or adjacent segments. Fig. 28. Sensory ganglion, human embryo, 20 weeks. Cells chosen are most advanced. 470 diameters. Fig. 29. Sensory ganglion cells, human embryo, 5 months. Cells chosen are most advanced. 470 diameters. Fig. 30. Sensory ganglion cells of adult dog. The nerve sfrom this ganglion had been stimulated by intermittent current of electricity for about ten minutes during anesthesia before death. The protoplasm is vacuolated, the nucleoli shriveled, the nuclei either swollen or shriveled, the pericellular lymph space is larger than normal, the tigroid masses have disappeared. 380 diameters. Fig. 31. Section through cervical enlargement of adult dog. Semidiagrammatic. Anterior aspect is upward. The posterior root fibers enter the cord and form synapses with the cells of the anterior horns. 10 diameters. The ganglion is enclosed within a tough connective tissue sheath which is continuous with the perineurium of the nerve trunk. The structure of the sensory ganglion is not so simple as was originally supposed. Two types of nerve cells are certainly known to be present in these ganglia, and others have been described by various authors. The first of these types is that which was formerly believed to make up the whole of the nervous portion of the ganglion. (Figs. 28, 29.) It has a large cell body with many fine dendrites and a single process which arises from an axon hillock. At a variable distance from the cell body, but within the ganglion, this process divides into two branches, one of which passes to the periphery and terminates in a sensory nerve ending, while the other proceeds to the cord and enters into various relationships to be described presently. These cell bodies lie within a pericellular sheath of connective tissue. A space is always to be found between the sheath and the cell body, which may be an artifact, or may be normally filled with lymph during life. This space is left larger than normal by the shrinking of cell body in fatigue or during certain abnormal conditions. (Fig. 30.) Another type of cell found in the spinal ganglion is that called after the name of Dogiel. It has many fine dendrites, and a short axon which breaks up into fibrillae within the ganglion. These fibrillae form basketlike networks around the other cell bodies of the ganglion, including those already described. These resemble in structure the Golgi cells of Type II, and probably they have similar functions. Amacrine cells also have been described in the spinal ganglia. Other cells have been described which have small cell bodies and relatively large nuclei which stain deeply. Thus they resemble embryonic cells. Nothing is known of their function. Some writers decribe non-medullated nerve fibers from these small cells entering with the posterior roots of the cord. The nerve trunks seem to consist chiefly of the processes of the group of cells first described. The posterior roots of the cord also are chiefly composed of the axons of these cells. In addition to these, the following structures have been described: I. Small medullated fibers, the axons of cells in the lateral horns of the cord, may pass out by way of the posterior roots, through the spinal ganglia, to enter the sympathetic ganglia. These are thus of the same class as similar fibers leaving the cord by its anterior roots. II. Non-medullated fibers from the sympathetic ganglia pass through the spinal ganglia to enter the spinal canal, and to be distributed to the vessels of the cord and meninges. III. Non-medullated fibers from the sympathetic system enter the spinal ganglia, to be distributed to its vessels. IV. Fibers have been described, upon rather doubtful evidence, which are from the sympathetic ganglia, which enter the spinal ganglia and enter into the formation of the pericellular baskets as in the case of the axons of the cells described in the second class. This relationship is not well studied as yet. The axons of the sensory neurons of the first order enter the cord as its posterior roots. (Fig. 31.) These fibers, upon entering the cord, divide into branches, one of which is short, and passes downward in the comma tract for the distance of one or two segments, while the other, which is longer, passes upward in the cord by way of the fasciculus gracilis (tract of Goll) to the nucleus gracilis, or by the fasciculus cuneatus (tract of Burdach) to the nucleus cuneatus. From both the ascending and the descending branches of the entering fibers collaterals are given off to the gray matter of the segment of the centering root, and to the segments immediately adjacent, both above and below. The destination of these collaterals is as follows: I. Collaterals form synapses with the cells of the anterior horns. In this way simple skeletal reflexes are governed. II. Collaterals form synapses with the cells in the posterior horns. In this way the more complex reflexes are governed, and certain classes of sensations are transmitted cephalad. III. Collaterals form synapses with the cells in the lateral horns. In this way the visceral reflexes are governed. IV. Collaterals form synapses with the cells in Clarkes column. In this way impulses are transmitted to the cerebellum, the delicate coordinations are secured, and the impulses of the pain and temperature are transmitted cephalad. The termination of the descending fiber is not known, but it is almost certain that it enters the gray matter, as do its collaterals. The ascending branches terminate, for the most part, in the nucleus gracilis and the nucleus cuneatus in the lower part of the medulla. A few enter the cerebellum by its inferior peduncles. The sensory neurons of the first order alone are immediately affected by changes in the environment or the condition of the body. Sensory neurons of the second order receive nerve impulses from the neurons of the first order, and transmit these impulses either to sensory neurons of the third and higher orders, or to associational or motor neurons. The termination of the fasciculus gracilis and fasciculus cuneatus in the nucleus gracilis and nucleus cuneatus in the lower part of the medulla has been mentioned. The axons of the cells in these nuclei carry the impulses cephalad by two paths. Fig. 32. Section through rhinencephalus of kitten. Cut freshened, with brain distorted so as to secure section of structures in horizontal plane. 5 diameters. Posterior ventricle; Horn of Ammos; Fornix; Optic thalamus; Candate nucleus; striae; Anterior Commissure; To temporal lobe; Olfactory tract; Frontal lobe. The larger number of these axons pass around the olive to enter the medial fillet, and thus pass upward through the formatio reticularis of the medulla and pons, into the tegmentum, and to the optic thalamus. They enter the thalamus and form synapses with the cells of the lateral nucleus of the thalamus. Just how many cells are intervened here is not known, but some of the cells of the thalamus send axons to the post-central convolution of the brain, where consciousness is affected. A smaller number of cells of the nucleus gracilis and nucleus cuneatus send axons into the cortex of the superior vermis of the cerebellum. The axons, probably of the Purkinje cells, transmit the impulses to the cells of the nucleus dentatus, from which they are again transmitted to the red nucleus, and from the red nucleus to the thalamus. From the thalamus the path is as before. Of the two ascending cerebellar tracts very little is known. It is true that the impulses concerned in the sense of muscular effort, perhaps including the impulses from the viscera, are carried to the cerebellum by way of the dorsal nucleus (Clarkes column) and the posterior ascending cerebellar tract (direct cerebellar tract, tract of Flechzig), of the same side, and in part by the anterior ascending cerebellar tract (tract of Gowers) of the opposite side. The muscular tone and the coordination of the muscular movements are probably governed in part through these impulses. From the cerebellum sensory impulses, probably of a very dull and indefinite nature, may be carried to the cerebral cortex and affect consciousness. The path of the impulses follows that already described. The impulses aroused by painful and thermic stimuli are probably carried by a devious way of the dorsal nucleus (Clarkes column), the anterior ascending cerebellar tract (Gowers) to the cortex of the superior vermis of the cerebellum and to the thalamus. The spino-thalamic tract diverges from the anterior cerebellar in the pons. From both these terminations the impulses are carried to the cortex by the paths already described. The axons of the terminal nuclei of the nerves of common sensation and taste in the medulla enter the opposite medial fillet, and are carried to the thalamus, and thence to the cortex. Other fibers from these terminal nuclei enter the cerebellum and form synapses with the cells, chiefly of the superior vermis. The axons of these cells transmit the impulses to the nucleus dentatus and by these cells again to the red nucleus, the thalamus and the cortex. Short paths may carry impulses upward and downward from one cell to another through the spinal cord, medulla, pons and midbrain. In cases of destructive lesions of certain fiber paths, it has been possible to secure a certain amount of sensation by education. In these cases the impulses travel extremely slow, and it seems probable that the short fiber paths, remnants of the primeval paths, have again assumed the duties which the higher development of the longer tracts had taken from them. The cranial nerves of common sensation are homologous with the spinal sensory nerves. Their ganglia are not to be distinguished from spinal ganglia by microscopic examination. The division of the processes within the ganglion, the amacrine and other cells of unknown function, the connective tissue network, all are present in the same relationship, so far as our present knowledge goes. The central prolongation or axon enters the medulla, divides into two branches, one of which passes downward, while the other passes upward, as do the spinal sensory roots. Both branches form synapses with the cells of the terminal nuclei of that particular nerve, and probably also of other nuclei of related function. The vagus has two ganglia, the jugular and the nodular (ganglion of the root and ganglion of the trunk of the vagus). The peripheral prolongations are distributed very widely, to the viscera of the throat, thorax and abdomen, to the ear, meninges, etc. The axons enter the posterior lateral sulcus of the medulla and form synapses with the cells of the ala cinerae, and probably also of the solitary nucleus. The glosso-pharyngeal nerve is concerned in the sense of taste Though a nerve of special sense, it has the structure of the nerves of common sensation. It has two ganglia, the superior and the petrosal, both placed at the jugular foramen. The axons enter the posterior lateral sulcus of the medulla and enter into synaptic relations with the cells of the solitary nucleus and the alae cinerae. The sensory root of the seventh, the intermediate, is also concerned in taste. Its axons enter the groove between the medulla and pons, and form synapses with the cells of the solitary nucleus and probably certain other centers. The trigeminal has a single ganglion, very large and distinctly tripartite. It is called semilunar, and lies near the apex of the petrous portion of the temporal bone. (It was formerly called the Gasserion ganglion.) It receives sensory impulses from the face, mouth, throat, nose, eye, ear, the anterior part of the scalp, and the meninges. Its axons enter the anterior face of the pons, and form synapses with a very long nucleus, reaching from the middle part of the pons, or higher, to the second cervical segment or lower. With the exception of taste, the neurons concerned in the special sensations enjoy certain marked peculiarities of structure. The nuclei of termination of the cranial nerves of general sensation and taste have the following relations: I. Axons of sensory neurons of the second order from these centers cross in the median raphe, enter the median fillet of the opposite side, and pass upward with that tract. Fibers are given off by this tract to both quadrigeminate bodies, to the red nucleus, substantia nigra, and to the nuclei of the reticular formation. The tract terminates in the lateral nucleus of the thalamus. From the thalamus the axons of the thalamic cells carry the impulses to the cerebral cortex. II. Axons enter the cerebellum chiefly by way of the restiform body. The impulses thus carried are concerned in coordination of the body movements, maintenance of muscular tone, etc. III. Axons pass to the various centers of the medulla. The impulses carried by these paths are concerned in modifying the activities of these centers in accordance with cranial sensory impulses. The acustic (auditory) nerve is made up of two parts, physiologically distinct, the cochlear and the vestibular. The ganglion of the cochlear portion of the nerve is the ganglion spirale. (Fig. 33.) It is placed in the modiolus, and follows the turns of the cochlea, whence its name. The dendrites of the cells pass to the organ of Corti and thus are affected by the sound waves. The axons of the cells enter the groove between the pons and medulla, plunge through the gray and white matter, and form synapses with the ventral and lateral auditory nuclei. Each fiber bifurcates near its nucleus, the two branches give off numerous collaterals, which form synapses with cells of the auditory nuclei.
Fig. 33. Ganglion spirale. Dark
areas are bones of cochleea. Embryo kitten, about 2 inches long.
40 diameters.
The axons of the cells of the ganglion spirale form synapses with the cells of two nuclei, the lateral auditory nucleus, placed on the outer aspect of the restiform body, and the ventral nucleus, placed in the medulla between the restiform body and the inferior olivary nucleus. A few fibers from both nuclei join the restiform body and enter the cerebellum. The axons of the lateral nucleus form most of the medullary striae. These fibers cross in the floor of the fourth ventricle, decussate, and plunge into the substance of the medulla to enter into the formation of the trapezoid body and the lateral fillet. A few of the fibers from the lateral nucleus pass with the fibers of the ventral nucleus. The axons of the ventral nucleus pass ventrally and turn toward the median raphe, where they decussate. Continuing toward the opposite side of the medulla they complete the formation called the trapezoid body. Among the fibers of the trapezoid body lie many small multipolar cells, with which the auditory fibers form synapses. The axons of these cells pass onward with the trapezoid body. The fibers turn abruptly forward on the side opposite their nuclei and form the lateral fillet. The bending of these fibers occurs at about the level of the superior olivary body. Many of the fibers terminate in the superior olive and in its accessory nuclei. The axons of the cells of these nuclei join the lateral fillet. The lateral fillet divides into two main bundles. Of these one enters the posterior quadrigeminate body of the same side. By this connection the reflexes governing the ear muscles are coordinated. The other bundle passes anteriorly by way of the inferior brachium to the medial geniculate body. From the medial geniculate body the axons arise, which enter the acustic radiations and carry the impulses concerned in the sense of hearing to the cortex of the first and second convolutions of the temporal lobe. A few fibers of the lateral fillet, either axons or their collaterals, enter the superior quadrigeminates. In this way the reflexes of the eye muscles following auditory stimuli seem to be coordinated. A few fibers enter the thalamus, and the impulses are then transmitted to the cortex with the acustic radiations. Other fibers seem to terminate in the nuclei of the reticular formation, and perhaps in the motor nuclei of the cranial nerves. The short peripheral fibers, or dendrites, of the vestibular ganglion (Scarpas ganglion) terminate among the hair cells of the cristae and the maculae. The axons enter the groove between the pons and medulla, and form synapses with the cells of the principal nucleus of the vestibular nerve (Dieters nucleus) and the nucleus of the descending root of the vestibular nerve. Each fiber bifurcates near its nucleus into an ascending and a descending branch. Each of these branches gives off numerous collaterals, which are distributed in the same manner as the terminals of the fibers. Some of the vestibular axons seem to enter the restiform body, and to be distributed with those fibers to the cortex of the superior vermis. The vestibular nerve is of interest in its phylogenetic development. Among the lower vertebrates the impulses from the middle ear and from the lateral line organs are of the most marked importance in adapting the movements of the fish to external environmental conditions. Among the animals of the upper vertebrate classes the lateral line organs disappear, the vestibular neurons are superseded in function in part by other more highly specialized neuron groups, and the vestibular nuclei, while retaining traces of their pristine relationships, have become almost reminiscent in function.
Fig. 38. Cross section through cord of
human embryo of about 10 weeks. Central canal is lined with ciliated
epithelium. Nerve cells are small, round, closely packed, with nuclei
occupying almost the entire cell.
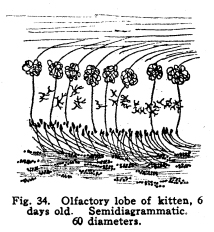
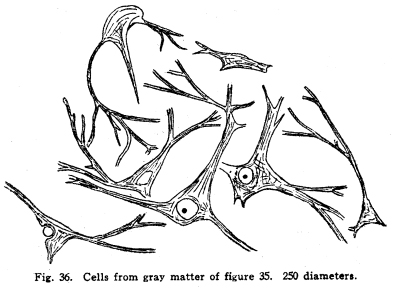
The chief vestibular nucleus lies beneath the medullary striae. Lateral to this lie the nucleus of Dieters and the nucleus of the descending root of the vestibular nerve. The nucleus of the descending root sends axons to the chief and to the accessory nuclei. Dieters nucleus receives fibers descending from the cerebellum, as well as the entering fibers of the vestibular nerve. The vestibular nuclei send axons as follows: I. Many fibers enter the cerebellum. Since the cerebellum is the most important center for the maintenance of equilibrium and for the coordination of the muscular movements, the impulses from the vestibule are of tremendous importance to that organ. Also, since the vestibular structures represent the lateral line sense organs, which originally were received by the cerebellar nuclei, the maintenance of the central relationships is very logical. II. Fibers decussate and enter the median fillet. With this tract they pass anteriorly, giving some fibers to both quadrigeminates, and to the nucleus of origin of the abducens nerve particularly, and to the nuclei of the reticular formation. They terminate in the lateral nucleus of the thalamus. The cortical connections are not well known. Probably the temporal lobe receives the impulses from the thalamus by way of the acustic radiations. III. Fibers from the vestibular nuiclei enter the nuclei of the motor cranial nerves. Various reflex movements of the cranial muscles are thus made possible. IV. Descending fibers terminate in the nuclei of the sensory cranial nerves, and in the nucleus gracilis and the nucleus cuneatus. V. Fibers pass to the inferior olivary nucleus; the function of this relationship is not known. It is probably concerned in muscular coordination and equilibrium. VI. Fibers pass downward through the cord as the vestibulo-spinal tract. This tract, of great value in fishes, is probably of very less importance in mammals. It carries impulses which are concerned in the maintenance of muscular tone, equilibrium, and perhaps in certain coordinations. Its injury is not productive of any but the most transient symptoms. The visual apparatus presents a number of variations from the typical sensory mechanism. In the first place, the retina is embryologically derived from a portion of the brain vesicles. It is not formed, as are the spinal ganglia, by wandering cells from the neural tube, but it is an outgrowth of the brain itself. Later, the part connecting the retina and the rest of the cerebral masses becomes atrophied. The axons of the ganglion cells of the retina follow the pathway of the obliterated nerve matter to the brain. The fibers of the optic nerves are, then, to be considered as homologous with the brain tracts, rather than with the roots of the spinal cord. In the retina several different types of neurons are found. The light first affects the layer of rods and cones, these transmit the stimulation to the layer of bipolar cells, these in turn affect the ganglionic cells, and it is the axons of these cells which transmit the nerve impulses toward the brain by way of the optic nerves and tracts. Among these nerve cells of the retina are found also certain othersthe so-called horizontal cells, whose axons pass horizontally and terminate among the bipolar and ganglionic cells, after the manner of the Dogiel cells in the sensory ganglia. Amacrine cells are found, whose dendrites branch freely among the other cell elements of the retina. There is some reason for supposing that the efferent fibers of the optic nerve terminate in the midst of the amacrine cells. If this be true, then the efferent impulses which, apparently, govern the nutrition of the retina, are effectual through the amacrine cells. There is room for doubt concerning the place of the visual sensory neurons of the first order. There is no doubt that the rods and cones are first affected by light, but the question arises concerning the nature of these structures. Their structure resembles greatly some of the specialized sensory nerve endings, and they are so considered by Barker and by certain other neurologists. The bipolar cells are thus to be considered as the sensory neurons of the first order. This view makes the bipolar cells homologous with the spinal sensory neurons, and the presence of the horizontal and amacrine cells in this layer of the retina adds favor to the view. The crux of the matter depends upon the real nature of the rods and conesif these be neurons, then the bipolar cells are sensory neurons of the second order. If the rods and cones are specialized sensory nerve endings, then the bipolar cells are of the first order, as Barker supposes. The matter is of importance only from the academic standpoint, and in considering the pathology of certain diseases of the nervous system. The ganglionic cells are homologous with cells of the nuclei of termination of the cranial sensory nerves. The optic nerves and tracts are homologous with fibers of the fillet. The fibers of the optic nerves and tracts are imbedded in neuroglia, as are the tracts of the central nervous system, and as true nerves are not. The embryonic development of the tracts follow the course of the development of the cerebral tracts. Just at the olivary body in front of the sella turcica the two optic nerves approach, and partially decussate. The fibers from the nasal halves of both retinae cross, while the fibers from the temporal halves of both retinae remain on the same side. The fibers emerging from the optic chiasm are called the optic tracts. The left optic tract thus contains the axons of the ganglionic cells of the left halves of both retinae, the right optic tract contains the axons of the ganglionic cells of the right halves of both retinae. The macula lutea is about equally represented by both crossed and uncrossed fibers. Of the optic tracts, about one-fifth of the fibers enter the superior colliculus and form synapses with the cells of its deeper layers. Here are coordinated the impulses concerned in the movements of the intrinsic and extrinsic eye muscles. The larger division of the optic tracts, comprising about four-fifths of the fibers, enters the lateral geniculate body and the pulvinar of the thalamus. It is not possible to determine how many cells are interposed in the pathway of the visual impulses through the thalamus, but certain of the axons finally pass to the cortex of the pole of the occipital lobe, where consciousness is affected. The bodies of the olfactory sensory cells lie in the mucous membrane of the nose. The region is called the regio olfactoria. It is very small, covering only about two hundred and fifty square millimeters on each side. These cells send dendrites toward the surface, where each fiber branches into very fine fibrillae, which are supported by modified epithelial cells called sustentacular. These fine branching fibrillae are capable of being stimulated by substances in gaseous form, or possibly also by substances in solution. The axons of the olfactory cells pass upward in twenty or thirty bundles through the cribriform plate of the ethmoid bone, and enter the olfactory lobes. These axons make up the olfactory nerves. In the olfactory lobe the axons pass around the periphery for a variable distance, then penetrate the lobe. Here they break up into fine branches and twist around into a globular form, making the olfactory glomerulus. Into the glomerulus also enter the dendrites from the mitral cells, in a deeper layer of the olfactory lobe. The axons of the mitral cells pass brainward as the olfactory tracts. (Fig. 34.) Fig. 34. Olfactory lobe of kitten, 6 days old. Semidiagrammatic. 60 diameters. Fig. 35. Spinal cord of shark. 10 diameters. Fig. 36. Cells from gray matter of figure 35. 250 diameters. Fig. 37. Section through cervical enlargement of adult dog. Semidiagrammatic anterior aspect is upward. The posterior root fibers enter the cord and form synapses with cells in anterior horn. The olfactory nerves are not medullated, as are all other cerebro-spinal nerves in the adult. The cell bodies lie in the mucous membrane. This arrangement differs from all other nerve cells of the body of vertebrates. Only in invertebrates are nerve cells to be found peripherally placed, with this exception. The axons of the olfactory neurons of the first order form synapses with the olfactory neurons of the second order by means of the glomeruli of the olfactory bulbs. The olfactory neurons of the second order, the mitral cells, and possibly also the granular cells of the olfactory bulbs, send axons brainward by way of the olfactory tracts. (Fig. 32.) The olfactory tracts carry fibers passing in both directions. Efferent fibers from the olfactory centers enter the olfactory lobes, and form synapses with the cells therein, apparently in all layers of true nerve cells. Efferent fibers seem to descend through the cribriform plate to the olfactory region of the nasal mucous membrane, though this matter requires further study. The olfactory tracts are composed of three roots. These, as they diverge, inclose the olfactory trigonum, an area of gray matter which is continuous with the anterior perforated space, the parolfactory area, the tuberculum, cinereum, and neighboring gray matter. The three roots are called lateral, medial and dorsal or middle. The lateral root passes directly to the amygdaloid nucleus, the hippocampus major, and neighboring areas. In passing, collaterals and terminals are given off to the gray matter of the inferior parts of the temporal lobe. Just how great an area of the inferior aspect of the brain and of the temporal lobe is included in the distribution of the lateral olfactory root is not yet known. It seems very probable, in the microsmatic human being, that many of the connections thus secured are reminiscent rather than actually functions. It is certainly true, however, that the function of the olfactory impulses in the unconscious coordinations is not to be measured exactly by any conscious factors, nor, indeed, in any exact manner by the phenomena of apparent reactions. The median root of the olfactory tract enters the anterior commissure. This requires a short discussion. This is a bundle of fibers which unite the lateral portions of the rhinencephalon. It is to be considered in two parts. The anterior part is composed of a bundle of fibers from the median olfactory root, which pass directly to the opposite side of the brain, and pass backward in the fornix, the stria medullaris and the septum pellucidum to be distributed, after losing some fibers in the gray matter traversed or passed, in the hippocampus major and the amygdaloid nucleus. The posterior bundle of the anterior commissure is composed of fibers which gather together from almost or quite all of the temporal lobes, decussate, and pass to the head of the caudate nucleus, to the areas of the temporal lobes, to the olfactory trigonum and the neighboring gray matter, and the olfactory bulbs, all of the opposite side. The anterior commissure pierces, or is pierced by, the stria medullaris and the external capsule fibers as they pass forward and medialward. It thus appears that the lateral roots are concerned in a rather scanty sort of crossing for the olfactory impulses. It is very evident that the olfactory impulses are largely carried by uncrossed tracts. The two sides of the rhinencephalon are intimately related by the various commissures. The dorsal or middle root of the olfactory tract plunges upward into the gray matter of the trigonum and neighboring gray matter. Part of the fibers form synapses with the cells in this area, but the larger number enter the fornix and the striae medullaris and pass to the hippocampus of the same side of the brain. Thus the fornix receives fibers of olfactory function at both its extremities, and the hippocampal region both receives and sends fibers by way of the fornix and striae. The hippocampal region of each side is related to its fellow of the opposite side by way of the horizontal fibers of the psalterium or lyre. This peculiar structure is concerned in carrying the olfactory impulses also. It is composed of the ascending and converging bundles of fornix fibers, between which pass small bundles of commissural fibers. These fibers carry impulses from each temporal lobe, and especially from each hippocampus, to the homologous contralateral areas. The further connections of the olfactory region are complex. The amygdaloid nucleus and the hippocampus, chiefly the uncus and dentate fascia, send axons by way of the fornix to the corpora mammillaria, partly of the opposite side, but chiefly of the same side. The fornix fibers terminate by forming synapses with the cells of the lateral or the median nucleus of the mammillary bodies. The axons of cells of the median nucleus of the mammillary bodies pass upward and bifurcate. The bundle which is composed of one set of these branches is the thalamo-mammillary bundle (bundle of Vicq dAzyr). It enters the anterior tubercle of the thalamus. Fig. 39. Sympathetic ganglion of adult dog. The pericellular baskets are shown. About 200 diameters. The second branch of the fibers from the median nucleus passes into the tegmentum and downward into the reticular formation. It seems to terminate in the neighborhood of the lateral nuclei of the reticular formation, probably in the superior and the central lateral nuclei. Collaterals and probably terminals are given off in passing to the red nucleus, substantia nigra, and hypothalamic nuclei. The axons of the cells of the smaller lateral nucleus of the mammillary body pass downward through the midbrain, and seem to end in the gray matter around the cerebral aqueduct. Fibers are given also to the red nucleus and neighboring centers. Through the mammillary bodies, the olfactory impulses are thus distributed to the centers which are concerned in the viscero-motor and the emotional and instinctive reactions. The nucleus habenulae also is concerned in the olfactory coordination. This nucleus receives the axons of the cells of the hippocampal region by way of the thalamic striae. These fibers enter the nucleus habenulae, both of the same and of the opposite sides, and form synapses with the cells therein. From the nucleus habernulae the fibers pass, as the fasciculus retroflexus (of Meynert), to the nucleus of the interpeduncular region, and the cells of this ganglion transmit the impulses thus carried to the nuclei of the motor cranial nerves. In this way the somato-motor effects of the olfactory impulses are carriedthat is to say, the olfacto-somatic reflexes are coordinated. |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}