
|
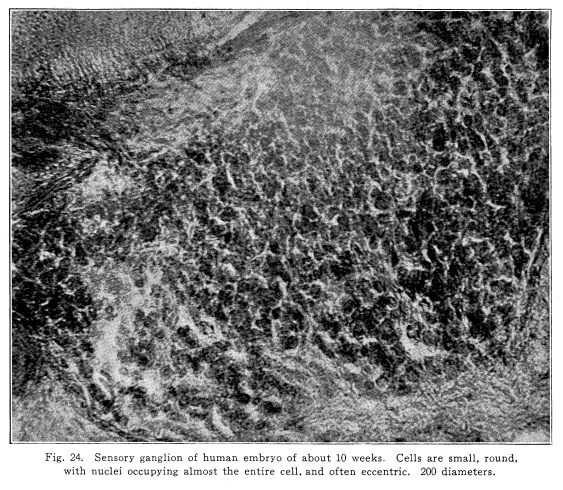
A nerve center is a group of nerve cells so related as to coordinate the nerve impulses which control any function. A nerve center may control any one organ, as the center of the sixth cranial nerve, or it may control a group of different organs, as the respiratory center, or it may control the functions of similar structures scattered over a wide area of the body, as the sweat center. Almost every imaginable degree of complexity of function and structure is illustrated in the nerve centers. All nerve centers have certain characteristics and all lie within the central nervous system. The nerve cells composing any center are very closely related in structure. This intimacy is accomplished in three different ways: I. Golgi cells of Type II are found. The short and freely branching axons of these cells connect cells in function which lie near one another. Amacrine cells may be found in some centers. II. Axons entering any centers, which bring to it the nerve impulses from other parts of the nervous system, give off collaterals to several cells of the center. Thus several cells are directly affected by the same entering stream of nerve impulses, and, through the mediation of the Golgi cells of Type II, this number is increased. III. The axons from the cells of the center give off collaterals which branch freely and form synapses with other cells of the center. Thus the activity of any cell may affect the activity of other cells of the same center. The number of cells thus directly affected is great, in some cases at least, but through the cells of Type II the activity of still other cells is affected by the same relationship. Centers which control the simple structures act by sending axons directly to the structures concerned in that function. Centers governing complex functions act by sending impulses to lower centers, to inferior and simpler cell groups which directly control the structures concerned in that function. Several different cell groups of varying degrees of complexity may thus be concerned in the government of certain complex coordinations. All centers are themselves subject to influence of other centers. We speak of higher centers, as if these were of authority. The term higher centers may be used in its structural sense, but only in a very restricted manner in its physiological sense. The centers which are higher are themselves as subject to the influence of impulses from the sensory and associational neurons as the lower centers are subject to the higher. In other words, the physiological activity of any center at any time depends upon the physiological condition of that center, and upon the algebraic sum of the nerve impulses which reach it from other parts of the nervous system. Ultimately, the source of all nerve impulses must be found in the incoming stream of sensory impulses which are derived from the changes in the conditions of the body to a certain degree, and to a greater degree from the changes in the environment of the body as a whole. The physiological condition of the cells of any center depends upon several factors. The nutrition of the nerve cell varies under physiological as well as pathological conditions. The food supply is in the lymph which bathes the cell. If the blood circulates too slowly, or if it contains less than the normal amount of nutritive substances, or if it is surcharged with the waste products of katabolism or with poisons either autogenic or otherwise, then the activity of the center is variously modified. In general, slight amounts of poison or fatigue, or slightly increasesd blood pressure, or temperature, increase the irritability of the nerve centers. Greatly increased pressure or temperature, or larger amount of poisons in the circulating lymph, lessen the irritability of the centers, and thus decrease their action. In the case of the respiratory and heart centers, probably also in the case of certain centers of the lower thoracic cord, the presence of varying amounts of carbon dioxid in the circulating blood is an important factor in controlling the physiological activities of the functions controlled by these centers. The physiological condition of any center depends in part upon its rest or fatigue. Nerve cells which have been resting, in which the tigroid masses are well formed and of angular outline, are normally irritable. If the cells have been active for so long a time as to cause a partial or complete disappearance of the tigroid substance, the activity of the center is lessened. The past experiences of any center modify its physiological condition. Each time any cell or cell group is stimulated the liminal value is lowered, and its irritability increased. This lowering of the liminal value seems to affect the cell for its lifetime. Thus, any center which frequently is called into action becomes constantly more readily stimulated. Any center which is subjected to excessive stimulation becomes abnormally irritable. If the stimulation is long continued, especially in an abnormal manner, the ultimate loss of function may result and may be followed by structural injury. This condition is exemplified in those cases of transverse myelitis due to excessive and abnormal sexual activity. The inactivity of any spinal center in the long-continued presence of bony malpositions or of the various peripheral irritations falls within this class of sources of abnormal action on the part of the nerve centers. The nervous system is supplied almost exclusively by terminal arteries. The occlusion of any one by pressure or by thrombus or embolus, or in any other way, thus results in the complete loss of nutrition in the part supplied by that artery. The area of infarction undergoes degeneration. The loss of the function of the nerve centers thus affected is absolute. Only the assumption of this function by other nerve groups can secure a certain amount of compensation for the injury. The nerve impulses reaching any center from other parts of the nervous system include impulses which stimulate the center to increased activity, and those which decrease its activity. These last are called inhibitory. The manner in which the inhibitory impulses act is not known, but it seems certain that there are impulses which do lessen the activity or the irritability of other nerve cells. Nerve centers act in accordance with the physiological condition of their constituent neurons. This is more easily seen in the autonomic centers. While the neuron is alive it, like other cells, is engaged in the processes both of anabolism and katabolism. During the period of inactivity the anabolic processes are more active than the katabolic. After a time, then, the accumulation of the anabolic products leads to an increased irritability of the neuron. This increased irritability may, in the absence of stimulation, become so great that extremely minute variations in the environment of the neuron, or perhaps even without such changes, the katabolic activities are initiated. This suggests the condition of affairs found when a child builds a tower with his blocks. He may place one above another, and another above that, and another, and another, until the instability becomes so great that the addition of a single block causes the destruction of the whole. Roughly, this may illustrate the phenomena of the action of the autonomic centers in the absence of stimulation reaching them. It must be remembered that this form of reaction is merely one of manythat the nerve centers rarely become so filled, or charged, with their anabolic products as to initiate the nerve impulse, because they are constantly subject, during anything like normal conditions, to the constant stream of stimulating impulses from other parts of the nervous system, and to the stimulation afforded by changing pressure of the blood and by changing characteristic of the blood itself. The typical nerve center receives axons from other nerve cells as follows: I. Collaterals from the axons of sensory neurons of the first order. By means of this relationship the center is enabled to act in accordance with immediate sensory impulses, and thus becomes a part of a simple reflex arc. II. Axons and collaterals from sensory neurons of the second order. By this connection more complex reactions are coordinated. III. Axons from other centers of related function. By this relationship it becomes possible for different centers to act synchronously, in a coordinated manner. IV. Axons and collaterals from the axons from the basal ganglia. By this means the impulses concerned in the various emotional reactions, and those reactions concerned with those simpler forms of emotions called feelings, are enabled to affect the action of the centers. V. Axons and collaterals from the axons from the cerebral cortex. This includes both the axons from the central convolutions, by means of which volitional actions are produced, and also the descending axons from other parts of the cerebral cortex, whose manner of action is not known, but which may be concerned in the phenomena of attention. VI. Axons and collaterals from the axons from the colliculi, from the pontine and olivary nuclei, from the vesticular nuclei, and from the cerebellum. The functions of these relations are not well known, but they may represent in part the relations important at an earlier stage of existence but now largely superseded by the development of the superior coordinations. The recognition of the physiological condition of centers which seem to be acting in an abnormal manner is a matter of considerable importance to the physician, and especially to the osteopath. For if any center acts improperly it is always because of some abnormal effect which is being produced upon it from without. As the centers do not act of their own volition, whimsically, during health, neither do they act abnormally without cause. It is not conceivable that any center should fail to act in a manner as nearly normal as its structure and surroundings permit. Nerve centers may act abnormally under the following conditions: The original structure may be such as to preclude normal activity. Efforts at compensation and adaptation probably form the sum of the possible therapeutics in such deformities. Sensory impulses of an excessive or deficient quantity may be sent into the center. The simple reflexes of which the center is capable may thus become either increased or decreased. If this condition continues very long, the liminal value of the neurons concerned may be so greatly lowered that later even normal stimulation may be followed by abnormal action. Visceral disease and subluxations of vertebrae and ribs are familiar examples of this condition. The impulses from the basal ganglia maya be affected by excessive emotional reactions, and the activity of the centers in connection therewith may thus become abnormal. This condition is especially true of hysterical subjects. In these patients there is rarely any consciousness of the real nature of the condition. Often these cases require the most careful study in their diagnosis. And the care of such patients depends altogether upon the recognition of the stream of abnormal impulses descending from the basal ganglia to the lower centers. Fig. 24. Sensory ganglion of human embryo of about 10 weeks. Cells are small, round, with nuclei occupying almost the entire cell, and often eccentric. 200 diameters. Nerve centers may become abnormal in function because of the abnormal quality or pressure of the blood circulating through them. The place of the vaso-motor impulses in governing the nutrition of the cerebral and spinal centers is not yet very well known, but there is no doubt that these are important factors in controlling the circulation through the nerve centers, as well as through other organs of the body. Whatever renders the vaso-motor impulses abnormal must, then, affect the nerve centers to some extent. It is essential for the normal activity of any nerve center that there shall be an unimpeded exit for the nerve impulses initiated by its activity. Rarely the activity of the lower centers may be rendered abnormal by abnormal volitional impulses from the cerebrum. Since the volitional impulses are by their very nature temporary in activity, and since the reflexes which control the lower centers are more powerful as well as being practically omnipresent, this source of malfunction of the lower centers must be seldom encountered, if ever. But the persistent abuse of the lower centers through overwork or poisoning or in other ways is responsible for much of suffering and disease. The
proper treatment in cases of abnormal action of the nerve centers depends
upon the recognition of the cause of the malfunction. In order to
secure the recovery it is only necessary to see that the cells affected
have rest if they need rest, exercise in due time and proportion, and the
unimpeded flow of good blood flowing rapidly under normal pressure.
Given these conditions, any nerve center not permanently injured must ultimately
become as nearly normal in function as its structural peculiarities permit.
In many instances the centers which have related functions are associated in the performance of those functions by means of neurons of association. The cardiac and vaso-motor centers, the cardiac and respiratory centers, are thus related. The nature of these relationships is not well known as yet. In some cases the connections are extremely complex, and in others it is only under decidedly unusual conditions that the physiological associations become recognizable. An examination of the sections taken from the medulla, for instance, shows each of the groups of nerve cells to be variously associated with other cell groups by means of axons and collaterals. It is true, also, that in other cases no association fibers can be demonstrated. It appears fairly evident that those centers which are most closely related structurally are, for the most part, those which have related functions. The development of the nerve centers is rather complex. The facts known are comparatively few, yet it is possible to use those few facts in explanation of certain relationships which otherwise appear meaningless. In the spinal cord of the lower vertebrates the gray matter is in great relative excess. With the higher development of the nervous system the process of cephalization is accompanied by the development of the long spinal tracts, and the gray matter thus becomes relatively less. In something of the same manner the process of cephalization leaves the centers of the medulla, pons, midbrain and cerebellum behind, with a relative diminishing of generalized functions and the assumption of more specific functions. In certain instances the lower center thus left behind becomes atrophied or lost. This is noted in the case of the vagal lobes, which are very important structures in the hind brain of fishes, but which are scarcely to be found in higher animals. In the process of cephalization the ascension of the functions of the centers and tracts is not ever complete. Probably always there are left behind a variable number of connections with the lower centers, and a number of the associational structures by means of which the lower centers were enabled to perform their duties. Thus, although the functions of the lower centers are more or less completely superseded by the development of the higher, with their more specialized structure and their more highly differentiated functions, the lower centers retain much of their pristine relationships. These structures usually assume other duties, sometimes subordinate to the higher centers, as in the case of the superior colliculi, sometimes of quite a different function, as in the case of the nucleus dentatus of the cerebellum. The complexity of the lower centers is more evident from the structural than from the functional standpoint. Many of the tracts of the medulla, pons and cord represent the structural relationships which are merely left behind in the process of cephalization, and which retain only a semblance of their one-time importance. The vestibulo-spinal tract, for example, was a tract of the greatest importance to the fishes; in mammals it is probably of the most minor import. In fishes it is concerned in the maintenance of equilibrium; in mammals it seems to be somewhat concerned with the maintenance of muscular tone, but its destructions by disease produces no perceptible symptoms. The midbrain shows plainly this process of developmental readjustment. The colliculi in fishes and amphibia are not differentiated plainly into superior and inferior. Almost the whole of the roof of the midbrain receives indiscriminately the visual, auditory and cutaneous senses. With the development of the centers of the thalamus, and still more of the cerebral hemispheres, these bodies assume different functions. The manner of their developmental changes can not be described here, partly for lack of space and partly because many of the steps are not yet well understood. It is enough for our present purpose that this development has occurred, that the colliculi become differentiated into superior and inferior bodies, that the superior developed the function of coordinating impulses to the eye muscles, while the inferior developed the function of coordinating the impulses to the ear muscles. The older functions of controlling the coordinations of practically the whole bodys movements, of being concerned in whatever dim sensations of sight the amphibia and fishes are capable of, have been assumed by the higher centers. Yet the colliculi still receive fibers from both the medial and the lateral lemniscus, and still send fibers downward through the cord as the tecto-spinal tract. The cerebellum was, in the beginning, simply the receiving center for the cutaneous sensations. With the disappearance of the lateral line organs, and with the development of the higher centers of the thalamus and the corpora striata, the functions of the cerebellum have been greatly modified. Yet the fibers carrying sensory impulses from the viscera, the somatic structures, and the sensory neurons of the second and higher orders still are carried to the cerebellum, and are of importance to it in the performance of its newer duties. The cephalization process is accompanied by the development of fiber tracts from the superseded centers to those of greater value in performing the duties. Thus, we have the complex by-paths of the sensory impulses by way of the anterior ascending cerebellar tract (Gowers tract), the brachium conjunctivum, the cerebellar cortex, the nucleus dentatus, the red nucleus, the thalamus and the cerebral cortex. This indirect route is explicable only upon the recognition of the process of the phylogenetic development of the cerebellar relationships. These instances are illustrative of the changes which have been recognized in the development of nearly all of the nerve centers of the medulla, pons and midbrain. The spinal centers seem to have been merely left behind in the race for supremacy; such functions as they retain show the original segmentation without such assumption of new functions as are to be seen in the centers named. Associated with the cephalization of the centers is the development of the long tracts of the spinal cord, medulla, pons, midbrain, and hemispheres. The connections between the lower and the higher centers thus become a matter of great convenience. These long tracts are the more subject to disease, partly, no doubt, because of their more recent phylogenetic development and their associated relative higher differentiation, and partly because of the fact that the longer tracts are made up of axons which are, because of their great length, separated from their cell bodies and the nuclei which control their metabolism. In the presence of abnormal conditions of the central nervous system the existence of the relationships phylogenetically ancient may be of considerable importance. We
live at this time in an age of great convenience. We talk by telephone,
send telegraph letters at night, expect to ride in airships, turn on light,
heat or water at will in our homes, speak to many people through printed
pages, and in many other ways we magnify our own personality by compelling
natural forces to be part of our means of expressing our own individuality.
Now, if any of us were placed in the forest, we could carry our own water,
cook food, build our own camping places, and manage to exist fairly well.
We could not send messages rapidly, but we should be glad of the pony express
if the wires were down and the railroads ruined.
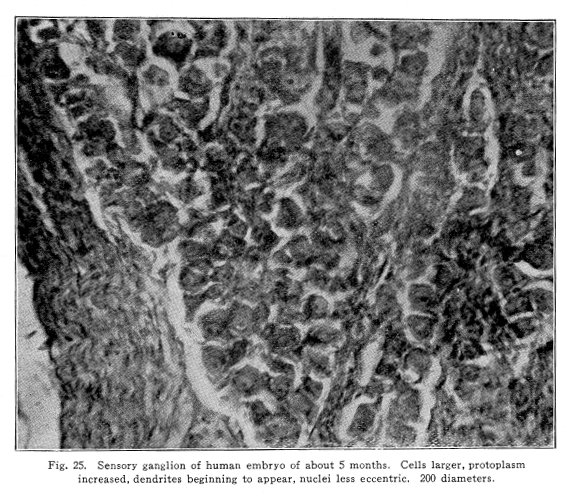
In cases of locomotor ataxis, for example, the degeneration of fasciculus gracilis, and sometimes part of the fasciculus cuneatus, is fairly complete. Yet the shorter tracts, carrying impulses of temperature, pain, and a certain amount of the tactile sense, remain for a long time in a fairly normal condition. These shorter tracts with their centers may be educated until they become functional in a degree sufficient to enable the patient to perform many acts not otherwise possible. Probably the speedy and efficient coordination of the normal nervous system can never appear, but the partial use of the lost functions is something. This education must proceed along the same lines as other methods of education. The liminal value of the centers yet active must be lowered by very carefully studied exercises. It must be remembered that overuse of any nerve center increases its liminal value, and that the same causes which led to the destruction of the longer tracts and the more highly developed centers may affect the shorter tracts and their centers also. So the exercises must be kept within the limits of fatigue always. The interest and attention of the patient secure the passage of the nerve impulses from the higher centers of the brain to the lower centers, and this stimulation lowers the liminal value of all the neurons concerned in the performance of the actions decided upon. The patient with locomotor ataxis may, for example, be set to use his feet in the normal position. The tendency is for the feet to be excessively rotated externally. Let him sit in his chair, pushing the feet forward with the toes straight forward, then withdraw them, watching them and guiding carefully every movement. Other exercises vary with individual patients. Thus the centers remaining normal may be educated to take up the duties of the injured centers and the short tracts may carry the impulses necessary for the performance of many duties. Such exercises may be used in many abnormal nervous conditions. This assumption of the duties long lost by the lower centers may interfere with diagnosis of cases of slow degeneration of certain nerve centers. This is evident in the light of experimental evidence. When the cerebellum, for example, is removed by several successive operations, the total loss of both hemispheres may be secured without the occurrence of any very noteworthy symptoms. This is, no doubt, due to the fact that the associated centers take up the work of the lost parts, and that they are able to do this because the slowly produced injury gives opportunity for the other centers to receive and send out the needful stimuli often enough to lower their liminal value, secure the permeable pathways in this way, and thus become functional. In case of diseases of the cerebellum, it may be that cases present no symptoms referable to the cerebellum at all, and only at autopsy is the real condition recognized. Probably in such cases the same assumption on the part of other centers through the slowly developing degeneration is responsible for the lack of symptoms. Similar conditions may be found in connection with all, or nearly all, of those centers which have undergone considerable changes in function during phylogenetic development. The condition is not usually true of those centers whose functions have remained unchanged during phylogenetic development, or in those whose functions are of comparatively recent origin. Even in the case of speech, however, the loss of the speech center on one side of the brain may be followed by the development of the speech center of the other side of the brain. In this case it is not a matter of the assumption of the ancient duties by the centers originally active in those duties, but it is the new development of the brain areas not before functional, either in ontogenetic or phylogenetic development. Fig. 25. Sensory ganglion of human embryo of about 5 months. Cells larger, protoplasm increased, dendrites beginning to appear, nuclei less eccentric. 200 diameters. |
{kind=link}
{kind=link}