
|
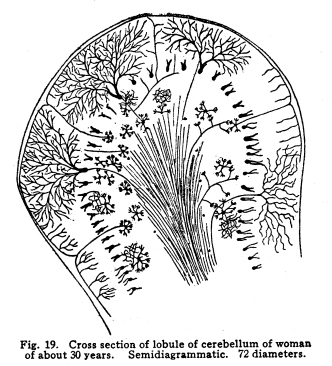
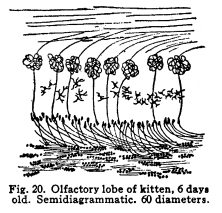
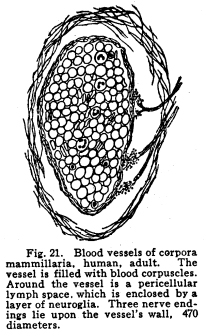
The individual neurons are functionally related by means of certain arrangements of dendrites, axons, collaterals and cell bodies, which bring the protoplasm of one neuron into very intimate structural relations with the protoplasm of another. This structural relationship which makes the functional relationship possible is called synapsis, from a Greek word which means clasping. Synapses are formed in various manners in different parts of the nervous system, and are often very complex. In all cases studied so far the axon or the collaterals from the axon of one neuron transmit the impulses to the dendrites or to the cell body of another. Thus, the axons are cellulifugal and the dendrites cellulipetal in function. The synapses of the neurons concerned in the more complex coordinations, especially in those controlling the autonomic functions, are so intimate, each neuron receives impulses from so many sources and sends impulses to so many other neurons, that the older views of Bethe and Golgi and Nissl, that the whole nervous system should be viewed rather as a syncytium than as a collection of individual cells, seems at first to be well founded. This appearance is noted with particular distinctness in the cell groups of the medulla, where are situated so many of the chief viscero-motor centers. Here may be well seen the extremely delicate fibrillae, resembling the extensions of the spongioplasm of the cell body into the pericellular space, which have aroused so great discussion. These fibrillae may be traced from one cell to another in so many cases and with such exactness that the existence of protoplasmic bridges can not longer be denied. Yet in these same centers may be found other cells, placed near each other, which seem to have no structural relationships. It seems probable that in the viscero-motor centers, at least, those neurons which are most closely related in function are those whose synapses are most intimate and complex. The synaptic relations of neurons are so varied that an almost infinite variety of reactions to any given environmental change becomes possible, as is, indeed, manifest to even the most superficial observation of the habits of the higher animals. Any one axon may give off a number of collaterals, all of which may form synapses with as many other nerve cells, and any one nerve cell may receive impulses from several different other nerve cells. When it is remembered that the nervous system contains thousands of millions of nerve cells, it is evident that the infinite complexity of even human thought and action does not surpass the complexity of the neuronic synapses. Probably the simplest form of synapsis is that found in the gray matter of the cord, as well as in other parts of the nervous system. The entering fibers of the posterior roots, the collaterals from the fibers of the spinal white matter, the axons of the Golgi cells of Type II of the cord, all break up into a very find feltwork which surrounds the intrinsic cells of the gray matter, the motor cells of the first orders and the cells of the viscero-motor centers of the cord. (Fig. 17.) The dendrites of the cells just mentioned branch freely among this complex feltwork, and by this means the impulses from several sources become able to affect the action of the muscles, the viscera, and the coordinating centers of the same and adjacent spinal segments. The Purkinje cells of the cerebellum display two very pretty methods of synapsis. The bodies of these cells are surrounded by a delicate network of collaterals from the axons of the stellate or basket cells of the cerebellum. This basket arrangement is similar to the feltwork around the spinal cells, but it is rather more easily visible and is more exactly limited to a single cell. (Fig. 18.) The dendrites of the Purkinje cells branch very freely in a manner resembling some old oak tree, except that the branches lie all in one plane. These dendrites are interlaced and wound around by fine vinelike climbing fibers, as they are called, the prolongations of cells whose location is not exactly known, but which almost certainly are carried into the cerebellum by way of the peduncles. (Fig. 19.) The olfactory lobes show another form of synapsis. The olfactory nerves entering the lobe pass tangentially over the surface for a variable distance. They then plunge into the mass of the lobe, giving off no collaterals; each axon branches and twines into a glomerulus. In the glomerulus also are found dendrites from the mitral cells of the olfactory lobe, which branch among the fibrillae of the olfactory axons. By means of this structure each cell of the olfactory region of the nasal mucous membrane is able to transmit impulses to a mitral cell, which, in turn, sends impulses brainward. (Fig. 20.) Fig. 19. Cross section of lobule of cerebellum of woman of about 30 years. Semidiagrammatic. 72 diameters. Fig. 20. Olfactory lobe of kitten, 6 days old. Semidiagrammatic. 60 diamaters. Fig. 21. Blood vessels of corpora mammillaria, human, adult. The vessel is filled with blood corpuscles. Around the vessel is a pericellular lymph space, which is enclosed by a layer of neuroglia. Three nerve endings lie upon the vessels wall. 470 diameters. Fig. 22. Prodasteroids from cortex of occipital lobe of new-born baby. 175 diameters. Fig. 23. Prodasteroids from medulla of adult cat. 60 diameters. In many parts of the nervous system are found cells, evidently nerve cells, which have freely branching dendrites, but for which no axons have been described. The function of these cells is not certainly known, but it seems probable that they are concerned in associating the nerve cells of their immediate neighborhood. Thus they would be similar to the Golgi cells of Type II in function. It may be, indeed, that the amacrine cells will be found to be of the same structure as the Golgi Type II cells. In the sympathetic ganglia each cell occupies a cagelike basket, which is formed of the interlaced fibers of the axons from the viscero-motor nuclei of the cord, medulla or midbrain. These small medullated fibers lose their sheaths near their termination and break up into the fibrillae which make up the pericellular baskets of the sympathetic ganglia. Any one basket may be composed of several axons or collaterals, and any given axon may send collaterals into several baskets. (I have seen five branches from a single fiber which was medullated before the branches were given off.) It seems probable, also, that any one fiber from the viscero-motor centers may pass through two or more sympathetic ganglia, giving off one or more collaterals in each ganglion. The synaptic relations of the viscero-motor neurons are thus no less complex in the ganglia than in the spinal and medullary centers. By means of the various methods of synapsis the neurons are able to affect the activities of one another and of the other parts of the body through the motor neurons of the first order. The nature of these effects are not yet well understood. It seems evident that neurons may affect other neurons in at least two different ways. Neurons may stimulate other neurons to increased action. This relationship is the most conspicuous. The stimulation of the sensory nerves may be followed by the passage of efferent impulses to the muscles innervated from the same spinal segment, or adjoining segments, or through the intermediation of the higher centers. Neurons may inhibit the action of other neurons. This relation is not easily understood, but is evidently one of the physiological facts. The stimulation of the cortex inhibits the activity of the spinal centers. This is noted in the excess of reflexes of any area associated with the injury of the centers or tracts above the spinal segments innervating that area. The inhibition of one neuron or neuron group by another is a very important phenomenon in nerve physiology, and it is of especial importance in the consideration of those centers associated with consciousness. A temporary inhibition of any nerve center gives time for the receipt of nerve impulses from other sources to affect the ultimate reaction. The nature of the inhibitions is not well known. There is some reason to assume a relationship between the phenomena of the refractory period and those of inhibition. In the case of mankind the simplest possible reaction is that of the simple spinal reflex action. In this arc we may consider the possible existence of a single sensory nerve fiber in relationship with the pericellular basket of a single motor cell. It is evident that the stimulation of the sensory neuron can affect the activity of that motor cell, and that only. It is not evident whether there might be at different times differences of the effect produced upon that cell by the receipt of the sensory impulsethat is, whether the physiological effects of the sensory stimulation might be subject to variation, as well as the liminal values of the two neurons concerned. The more complex spinal reflexes depend upon the coordinate action of several neuron groups. It is evident that the interpolation of even a very few relay stations adds very greatly to the structural possibilities of the reaction which might follow upon the receipt of the sensory stimulus. Probably the simplest action of which we are capable, even reflexly, necessitates the activity of many nerve cells, extending through several spinal segments. Another more complex arc includes in its circuit the spinal efferent and afferent cells, and the cerebellar centers. Reactions which are subject to the cerebellar coordinations are as complex as the habits which we learn, and which become as fixed as any reflex action whose foundations were laid in the nervous system long before birth. Yet more complex are the arcs which include the cells of the centers lying about the base of the brain, in which the emotional and instinctive reactions are coordinated. In these centers are received, correlated, and sent out again, the impulses concerned in the movement not only of the skeletal muscles, but also of those visceral activities which have been found associated with the best and longest life of the individual and his race through all the phylogenetic developmental steps. Here are related in function the impulses concerned in the erection of the hair in fright, the grinding of the teeth in anger, the movements of the nose in disgust, and all the other phenomena of the instinctive and emotional reactions. These various reflex arcs are complex; their action necessitates many coordinate reactions on the part of millions of nerve cells; their activities are the result of ages of inheritance of those whose nervous systems were adapted to performing their duties most efficiently, and through the working out of the many other laws which govern racial development. Yet all of these reactions appear very simple and predictable when the arcs which include the cerebral cortex are noted. Receiving impulses from all parts of the body, the cortical centers coordinate and control the impulses arising from the activities of the lower centers. The impulses are also coordinated in relation to the past effects of previous experiences. Through the intermediation of consciousness, not only the actual past may be enabled to affect the ultimate reaction, but the elements of past experiences may be dissociated and recombined, in order that the ultimate reaction may be wise beyond experienceor foolish beyond experience, if the dissociations and the recombinations were ill advised. All of this multiplicity of reaction capacity and reaction choice becomes possible only through a multiplicity of synaptic relationships. In any given neuron group, or neuron system, it appears that there must be an element of choicenot, of course, necessarily conscious. If any one neuron in synaptic relationship with two other neurons should be stimulated, it is evident that three different reactions are possible, excluding qualitative differences. The stimulation of the first may cause the stimulation of either of the two to which it is related, or it may cause the stimulation of both. If one neuron is associated with three or more others in function, the number of the possible reactions becomes increased to seventhe stimulation of any one of the three, of any combination of two out of the three, or of all three of the neurons. It is evident, then, that the element of physiological choice becomes a matter of some complexity. So far as these relationships have been studied, the factors which determine the choice in any given case are as follows: I. The impulses from any nerve cell are apt to be carried over the axon rather than over the collaterals from the axon. Thus, the impulses from the motor neurons of the cortex are carried directly to the lower motor centers, without affecting the basal ganglia cells, unless the stimulation of the cortical cells be profound. In case of increased stimulation, the impulses are carried over the collaterals also, and, in most cases, more active movements result than is the case of the lighter stimulation. II. Of two neurons in synaptic relationship with a third, that one which has the lower liminal value is the more apt to be stimulated. The factors which modify the liminal value of any neuron have already been discussed. III. There is some evidence in favor of the view that the gemmules upon the dendrites of the nerve cells are capable of amoeboid movement. If this be true, it is possible that the retraction of the gemmules or of the dendrites may modify the receptivity of any neuron. IV. There is, in nerve cells as well as in muscle, gland, and other active cells, a certain refractory period following stimulation, during which any stimulus received is followed by no perceptible effect. This refractory period is very short in the case of the nerve cell, but is demonstrable in so many instances that it is fair to assume it to be a characteristic of neuron physiology in general. If, of two neurons receiving impulses from a third, one has been so recently stimulated that the refractory period is not yet passed, the other is the more apt to be affected. V. Impulses which arise from those parts of the body nearest the spinal cord are most apt to initiate deep-seated reflexes than impulses arising from structures more distant from the cord. This is doubtless due to the fact that the innervation of the body is determined at a time when the cord occupied a position somewhat more nearly the center of the structures innervated. The budding of the limbs and the changing form of the thorax and the abdomen modify the areas of innervation, and while the number of nerve cells and fibers becomes increased to a certain degree, the central relationships do not appear to be rendered more complex in correspondingly great degree. Thus we have the malpositions of the vertebrae, a source of greater ills than malpositions of the bones of the hands, for example. Irritation of the skin over the back of the neck may be a source of considerable reflex irritation, but the irritation of the skin over the arm or the foot has much less of an evil effect, so far as the deep reflexes are concerned. VI. Sensory impulses are more efficient in arousing any given reflex action the more nearly the functional relation between the area stimulated and the area affected by the efferent impulses. The sensory impulses from any part of the naso-pharynx, for example, are more apt to affect the other parts of the respiratory tract, producing a sneeze or a cough, than to affect the activity of the muscles of the pharynx, which are nearer anatomically but less closely related in function. Sensory impulses form the buccal pharynx, on the other hand, are more apt to initiate the reflex actions concerned in vomiting. VII. Those impulses affect consciousness most vividly which arise from parts of the body most immediately affected by environmental changes. Impulses from the skin are clearly perceived in consciousness and are located with more or less accuracy. Impulses from those parts of the skin most subject to stimulation are those in which the localizing sense is most acute. Impulses from the skin of the back, for example, while fairly well adapted to the production of reflex effects, are not well localized in consciousness. VIII. Other things being equal, those impulses arising from parts of the body whose nerve centers are being left behind in the process of cephalization, or which are themselves in process of phylogenetic regression, are least apt to affect consciousness, and are most apt to be efficient stimuli of the reflex activities. This is noticed in the case of the vestibular sensations, which are scarcely to be recognized in consciousness even when the attention is given to the effort, yet which are very efficient in arousing reflex actions. The phenomena of seasickness probably illustrates this reaction, and also the phenomena of Menieres disease The centers for the area supplied by the lower sacral nerves seem to be in process of regression. The impulses from this part of the body are not especially well adapted to affecting consciousness, though injuries of them may cause extreme suffering, but even slight injuries of this area are often associated with reflex effects out of all apparent proportion to the sensory disturbance. IX. Conscious attention to any sensory impulses increases the power of those impulses to affect consciousness. This phenomenon may add to the suffering under certain conditions of disease. Other factors not yet studied are probably responsible for many of the inexplicable effects of stimulation. In the case of the peripheral irritations, for example, it is impossible to determine what effect will be produced in any given individual. The nature of the relations which make possible the inhibition of one neuron by another needs more study before our knowledge of the interneuronic relations is at all satisfactory. That these relations are capable of considerable modification seems probable from facts observed in certain clinic cases. The effects of cross suturing of nerves, both experimentally upon animals and in traumatic paralysis in man, seem to indicate that the permeable pathways of impulses through the central nervous system are capable of change. In locomotor ataxia, also, the results of careful re-education show that probably nerve impulses may ultimately employ pathways which, under normal conditions, would not be used. Many of these things are not to be explained satisfactorily with our present knowledge of the functional relations of the neurons. |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}