
|
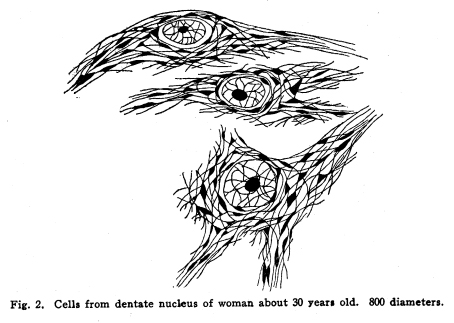
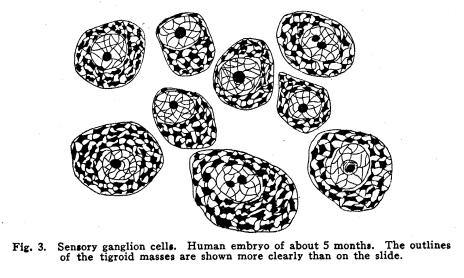
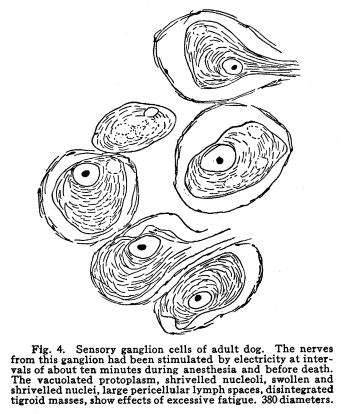
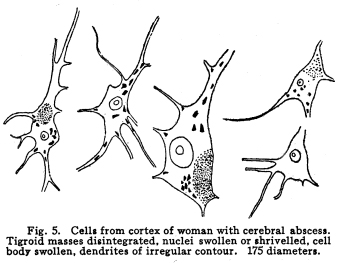
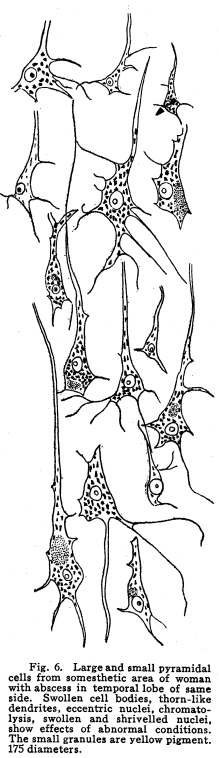
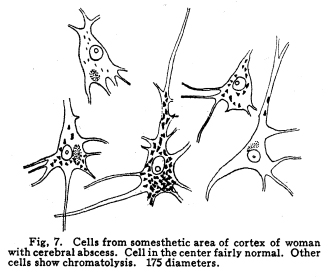
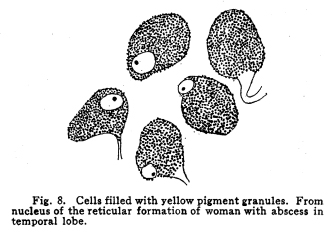
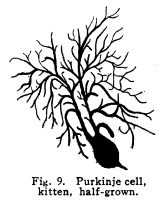
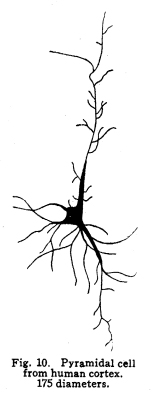
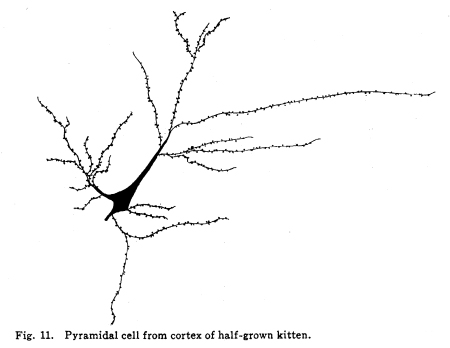
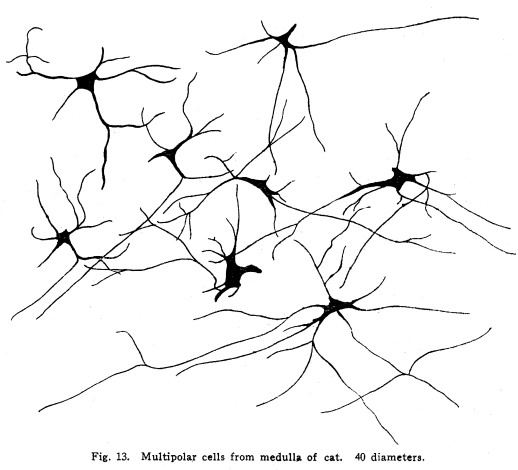
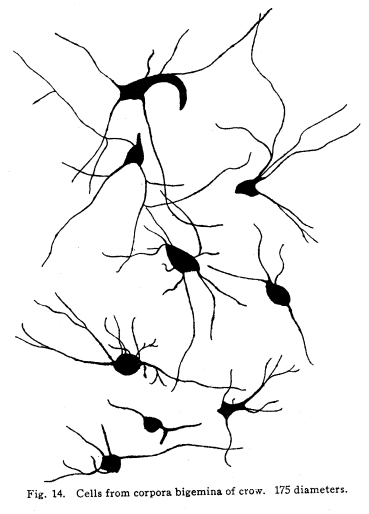
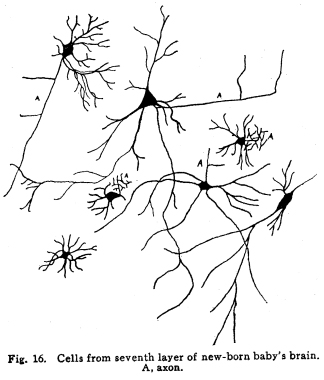
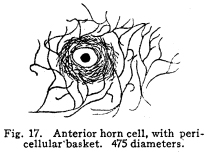
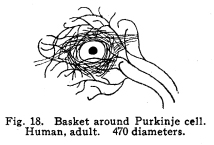
The nervous system has been grossly divided into various parts, such as cerebrum, cerebellum, spinal cord, ganglia, and so on. This gross division is convenient for purposes of study, but it must be remembered that it does not represent any logical classification of the parts of the nervous system. Any separation of one of these parts from its fellows can be accomplished only by cutting through numbers of fibers which belong to the cell bodies of other and often distant parts of the nervous system. The units of which the nervous system is composed are neurons and the tissues which nourish and support them. The unity and independence of the neuron has been demonstrated. This is called the Neuron Theory, that is, that the neuron is structurally and physiologically an individual, preserving its identity throughout life. It is not capable of independent existence. It has, after a very early period of embryonic life, no power of reproduction; throughout life it requires for its nutrition very complex substances which must be formed by other tissues of the body. It is thus an extremely specialized cell, though it is as independent, as individual, as much a structural and functional unit as is the cell of the liver or of the blood. It seems also true that each neuron, or at least each group of neurons, is specialized, doing its own work and none other; unable to perform the duties of other neurons, as they are unable to perform its duties. As the death of persons of unusual ability leave work long undone, so the injury or death of these highly specialized cells leave undone those duties for which they were especially adapted. The external form of neurons varies greatly. (Fig. 1.) The embryonic cells are spherical. By the outgrowths of the axon and dendrites the shape assumes many variations, many of them very complicated. The processes of the neuron include dendrites and axon. Within the protoplasm lie various deutoplasmic substances. The nucleus, in the adult normal neuron, lies near the center of the cell body. All of these structures, while in the main resembling similar structures of other cells, yet present certain peculiarities. The nucleus varies in size both absolutely and relatively to the size of the cell body. Throughout the nervous system two chief classes of cells are to be found, depending upon the size of the nucleus and the cell body. This classification is given by Nissl. Cells whose nuclei are large, lying within a scanty ring of protoplasm, are called karyochromes. They resemble embryonic cells, though they may be found during life. The karyochromes have deeply staining nuclei, with the nucleoli rather poorly defined. The protoplasm is very scanty, contains no tigroid masses or pigment, and has no very well defined reticulum. The nucleus may be eccentric. The cell has no well defined axon or dendrites, but may have one or two short prolongations, not to be distinguished as either axon or dendrite. The function of the karyochromes is not known. Somatochromess are of larger size, with nuclei relatively small, lying in the midst of a large mass of protoplasm. Probably the functional part of the nervous system is composed of somatochromes. The somatochromes have nuclei which stain feebly and with difficulty. Their nucleoli stain deeply and are well developed. Both the nuclear and the cytoplasmic reticulum are well defined. The protoplasm surrounds the nucleus about equally on all sides in normal cells. The axons and dendrites are variously developed, becoming of great length in some cases. The cell body and the dendrites contain tigroid masses and pigments in amounts varying with the class of the cell, its place in the nervous system, and its physiological conditions, such as rest, nutrition, age, etc. The nucleus is permeated by a chromatin network, with knots at the intersections. This network contains within its meshes one nucleolus which stains very deeply and is alwaysor seems to be alwayspresent. Besides this nucleolus there may be from one to four others, which are sometimes called supernumerary. These stain less deeply and often variably, and they may vary greatly in size as well as in numbers. These supernumerary nucleoli may be simply rather large net-nots. The centrosome is not found in the neuron after it has reached a stage of development sufficient to render its recognition possiblethat is, after it has passed the possibility of reproduction. A few observers have reported centrosomes in the nerve cells of adult brains in the neighborhood of injuries, but these findings have not been substantiated by later studies. It now seems probably true that the later divisions of the neuroblasts were of such a nature as to give the structures concerned in initiating division to the cells which become developed into neuroglia, while the cells which become developed into neurons are thus left without the power of reproduction but with enormously increased possibilities of differentiation along the lines of irritability and conductivity. Later investigators report the centrosome present in sympathetic cells at a later time. The function of the nucleus in the neuron is of the same nature as in other cells. It controls the nutrition of the neuron throughout its whole extent. In the case of the cells of the sensory ganglia of the lumber cord, the peripheral prolongation may be, in a tall man, more than a meter in length, while the centrally directed axon is about equal in length. The nucleus of these cells, placed about midway of the whole length of the neuron, controls the nutrition of the whole. The cytoplasm of the neuron is of extreme complexity. The spongioplasm is composed of rows of granules which stain with varying degrees of intensity. The fibrils of the spongiaplasm extend into the dendrites and beyond the limits of the hyaloplasm, and similar fibrils extend into the axon. The axonic fibrils stain in a manner slightly different, but by some neurologists are considered continuous with the spongioplasm of the cytoplasm. In the meshes of the spongioplasm lie the hyaloplasm and certain deutoplasmic bodies. The hyaloplasm is a homogeneous, viscid substance, which stains very feebly with the protoplasmic dyes. Very little is known of its structure or function. The tigroid masses, or Nissls substance, or chromophilic granules, as they are variously called, lie in the meshes of the spongioplasm of the cell body and the proximal part of the dendrites, but not in the axon or in the axon hillock. (Figs. 1, 2, 3.) These bodies are usually found only when the nerve cells have been fixed very rapidly and stained in a certain manner. Intravitam staining does not demonstrate them clearly, though sometimes they may appear faintly. For this reason it is supposed that they do not exist as such during life, but that the appearance is a post-mortem phenomenon. It is, however, a very constant and valuable phenomenon, since the appearance of the tigroid substance changes very profoundly during fatigue, poisoning, or disease. The tigroid substance differs chemically from the other neuron structures. It is not soluble in dilute and concentrated acids, boiling alcohol, cold or boiling ether, or chloroform, and it resists the action of pepsin-hydrochloric-acid longer than do other cell structures. It can be dissolved from the cell, leaving the cytoplasm intact, by the use of concentrated solutions of lithium carbonate. These masses contain iron, phosphorus and the nuclein bases in about the proportions of nuclear material. Fig. 1. Cell from hippocampus of kitten. 100 diameters. Fig. 2. Cells from dentate nucleus of woman about 30 years old. 800 diameters. Fig. 3. Sensory ganglion cells. Human embryo of about 5 months. The outlines of the tigroid masses are shown more clearly than on the slide. The masses are composed of fine granules imbedded in some coagulum-like material. They are usually angular in outline in the normal adult cell. Similar chromophilic masses are to be found in the nerve cells of invertebrates, even those of rather a low type, such as snails, molluscs, etc. In abnormal neurons, those fatigued, poisoned, or affected by certain conditions of disease, the tigroid substance shows very pronounced changes from the normal. (Figs. 4, 5, 6, 7.) At an early stage of fatigue the masses are found to have rather rounded outlines and to stain less deeply. Later they are found smaller, with even less vigor in staining. With exhaustion, the masses are not to be found, and the cytoplasm of the nerve cells takes a faint blue tint, with perhaps here and there very fine particles with the deeper stain. Under the influence of poisons, excessive heat and certain diseases the variations in the size and staining of the tigroid masses are very characteristic. This matter is discussed at length in Barkers Nervous System. Since the tigroid substance shows these constant changes, since it becomes dissipated during cell activity and becomes restored during rest, it is supposed that it represents the reserve of cell energy. It is not known whether the disappearance of the tigroid substance is due merely to a mechanical separation of its particles, or whether it is actually used up during cell activity. Since the masses are rebuilt in a remarkably short time, it must be true either that they are simply separated during activity and reunited during rest, or that the materials of which they are composed are very rapidly built up from the lymph surrounding the nerve cell. In certain tests they have been found to be restored to their normal appearance within twenty minutes. Fig. 4. Sensory ganglion cells of adult dog. The nerves from this ganglion had been stimulated by electricity at intervals of about ten minutes during anesthesia and before death. The vacuolated protoplasm, shriveled nucleoli, swollen and shriveled nuclei, large pericellular lymph spaces, disintegrated tigroid masses, show effects of excessive fatigue. 380 diameters. Fig. 5. Cells from cortex of woman with cerebral abscess. Tigroid masses disintegrated, nuclei swollen or shriveled, cell body swollen, dendrites of irregular contour. 175 diameters. Fig. 6. Large and small pyramidal cells from somesthetic area of woman with abscess in temporal lobe of same side. Swollen cell bodies, thorn-like dendrites, eccentric nuclei, chromatolysis, swollen and shriveled nuclei, show effects of abnormal conditions. The small granules are yellow pigment. 175 diameters. The tigroid masses persist in remarkable manner during certain disease processes. In a section from a brain containing an encysted bullet and an abscess, together with numerous small foci of infection, there are often found cells closely adjacent to the abscess and the inflamed areas, and to the bullet cyst, which contain many fairly normal tigroid masses, while in the same section, not more than the diameter of the cell distant from those with normal tigroid substance, are found others with every appearance of severe degeneration, and with a total absence of the tigroid masses. The yellow pigment is another of the dentoplasmic substances of the neuron. This substance occupies certain restricted areas in the cell body. (Figs. 5, 6, 7.) The pigment is composed of rather coarse granules, which are of a light yellow color. It is not dissolved by ether, oil, alcohol, or water; it is not affected by any of the dyes usually used in preparing neurological material. It is stained with osmic acid if it has not been acted upon by ether or alcohol. It is not identical with the pigments of substantia nigra, locus ceruleus, etc. This pigment is not found in embryonic material. In the human nervous system it is first found in the spinal ganglion cells of the child of about six years of age. At nine it is found in the motor cells of the cord. Later it is found in the brain and all through the central nervous system. It is not described as occurring in the cells of the sympathetic system. Under abnormal conditions, as in general paresis, or in premature senility, or in the presence of brain lesions, the amount of the pigment may be enormously increased. It may occupy practically the whole of the cell body, leaving the nucleus half extruded. Sometimes the cell leaves no trace of its existence save that the mass of yellow pigment shows the outline of the cell as it appeared during its life. (Fig. 8.) Fig. 7. Cells from somesthetic area of cortex of woman with cerebral abscess. Fig. 8. Cells filled with yellow pigment granules. From nucleus of the reticular formation of woman with abscess in temporal lobe. Fig. 9. Purkinje cell, kitten, half-grown. Fig. 10. Pyramidal cell from human cortex. 175 diameters. Fig. 11. Pyramidal cell from cortex of half-grown kitten. The facts known in regard to the yellow pigment present this substance to us as if it were an insoluble waste product of nerve metabolism. If there were some substance, insoluble in the fluids of the body, and formed in very small amounts during a lifetime, such a substance would vary in amount and distribution as the yellow pigment varies. Other pigments are found in the bodies of the neurons of the substantia nigra, the locus ceruleus, etc., but these seem to be constant during extra-embryonic life. The dendrites, as their name indicates, resemble trees. This resemblance is very beautifully shown in the Purkinje cells of the cerebellum. (Fig. 9.) They are protoplasmic prolongations of the cell body, and they have the same staining reactions as the protoplasm of the cell body itself. These branches originate as outgrowths of the cell body. They have broad bases and usually lose in diameter during their length. (Fig. 10.) They branch at acute angles, like the branches of trees, and they exhaust themselves sometimes through frequent branchings. They are usually quite short, and do not leave the vicinity of the cell body. In the case of the sensory neurons of the first order, however, the dendrites attain enormous length, assume a medullary sheath and neurilemma, and are not to be differentiated, structurally and in adult life, from axons. Fig. 12. Cell from hippocampus of kitten. The cortex had been stimulated by electricity during anesthesia before death. 100 diameters. Fig. 13. Multipolar cells from medulla of cat. 40 diameters. Fig. 14. Cells from corpora bigemina of crow. 175 diameters. Fig. 15. Polymorphic cells from medulla of adult guinea pig. Fig. 16. Cells from seventh layer of new-born babys brain. A, axon. Fig. 17. Anterior horn cell, with peri-cellular basket. 475 diameters. Fig. 18. Basket around Purkinje cell. Human, adult. 470 diameters. The dendrites within the central nervous system are often found studded with small budlike protrusions called gemmules. (Fig. 11.) It is not certainly known whether these gemmules are an artefact or whether they represent a normal structure of the neuron. In certain diseases of the nervous system, particularly the brain, these gemmules are found greatly swollen and of irregular outline and position. (Fig. 12.) Sometimes they do not appear at all in normal brain material; sometimes they are found plainly in equally normal material. Their nature is thus unknown at present. Dendrites contain the tigroid substance, as does the body of the nerve cell. Dendrites rarely contain the yellow pigment granules. The function of dendrites is not certainly known. There is some reason for supposing them to be partly nutritional in function. Nerve cells are unusually large. The surface of the cell body, that is, the possibility of absorbing nutrition and of excreting wastes, is proportionately small in comparison with the mass of the cell, that is, with its need of nutrition and its formation of waste material. This relation seems the more striking and fatal when it is remembered that the metabolism of the neuron is of an extremely rapid and vigorous order, and that its activity is so complex that there is great need for the most speedy renewal of the nutrition and most speedy removal of its wastes. Now by means of the forest of finely-branching dendrites the total surface area of the cell body is greatly increased, and the facilities for the absorption of nutrition and for the removal of wastes is correspondingly increased. There seems very little doubt that this is one important function of the dendrites of the cell. (Figs. 13, 14, 15.) Dendrites are known also to carry cellulipetal impulses. Morat insists upon the polarity of the neuronthe fact the impulses reach the cell body by means of the dendrites and leave it by means of the axon. This polarity is very well demonstrated in the case of certain neurons. An important structure of the neuron is the axon. It is probable that the neurons of the higher vertebrates, at any rate, possess only one true axon, though the sensory neurons of the first order have two processes which are very much alike. Physiologically, however, even these processes differ, since the peripheral prolongation carries cellulipetal impulses; and this process contains the tigroid substance during its early development. In other parts of the nervous system the mon-axonic nature of the neuron is evident. The axon arises from a part of the cell body which contains no tigroid substance. (Fig. 6.) During embryonic development and throughout life the absence of the tigroid substance in the axon and in the neighboring protoplasm is constant. This space around the origin of the axon is called the axon hillock or implantation cone. It may be placed almost anywhere upon the surface of the cell body, or upon any of the larger dendrites near the cell body. (Fig. 16.) The axon retains its diameter throughout almost or quite its entire length. It may give off branches called collaterals, which arise at a right angle to the axon, or may assume a somewhat recurrent direction. Neurons are classified by Golgi according to the form of the axon. Cells whose axon is long, and passes into the white matter, are called by him Type I cells, while those whose axons are short, giving off many very short collaterals which ramify extensively in the immediate neighborhood of the cell body, are called Type II cells. It is evident that Type I cells are concerned in relating parts of the nervous system which are more or less distant from one another, while the Type II cells bring into relation those neurons which are placed very near one another. The spongioplasm of the axon and the axon hillock seems to be continuous with the spongioplasm of the cell body itself. The axon contains fibrils which differ somewhat from those of the cell body and dendrites in staining reactions, though they seem to be more or less continuous with them. Axons vary in length from the few microns of the Golgi cell of Type II to the meter or more of the axons of the large cells in the anterior horn of the lumbar cord, which terminate in the muscles of the feet, or the axons of the lumbar sensory ganglion cells, which terminate in the nucleus gracilis in the medulla oblongata. The nutrition of the axon, in all its extreme and attenuated length, is dependent upon the integrity of the neuron as a unit. As in every other cell, once any part of the protoplasm is severed from the nucleus, that part soon becomes degenerated and dead. In the case of the long axons the mass of the fiber may be two hundred times the mass of the cell body, and yet the small cell body with its nucleus controls the metabolism of the axon to its farthest extremity. The axon carries cellulifugal impulses, that is, it carries the nerve impulses from the cell body to other neurons, or to the axon terminations in muscles, glands, etc. Long axons in the central nervous system are surrounded by the myelin sheath, or medullary sheath, or the white substance of Schwann, as it is variously called. This is a white, homogeneous substance of a fatty nature, and it surrounds the axon in sufficient quantities to make the area of a cross section of the sheath equal to the area of the cross section of the axon which it encloses. This myelin substance gives the characteristic white glistening appearance of the so-called white matter of the brain and spinal cord. The olfactory axons alone of the cerebro-spinal nerves are non-medullated. Axons become medullated in the order of their functional development, though it is not known whether beginning function precedes or follows the medullation of the axons. The axons of the sympathetic neurons have either no medullary sheath or extremely thin ones, in mammals. In birds the sympathetic axons are usually medullated. Outside of the central nervous system the axon has another coat, the neurilemma. This is a sheath composed of connective tissue cells, greatly flattened and applied very closely to the medullary sheath. At intervals of about seventy-five times the diameter of the nerve fiber the medullary sheath of the peripheral nerves is interrupted by a circular constriction which permits the neurilemma to lie in contact with the axon itself. These interruptions are called the nodes of Ranvier. The neurilemmal sheath between one node of Ranvier and the next contains one nucleus, and is thus supposed to be derived from a single connective tissue cell. The function of the medullary or myelin sheath is not known. It has been supposed to act as a sort of insulator, as a protection to the nerve fiber, as a source of nutrition to the nerve fiber, but there is no sufficient evidence in favor of any of these views. The neuron is a cell and has the structure characteristic of all cells. It is also a highly differentiated cell, and has, in addition to these, the structures adapted to the performance of these specialized functions. The neuron consists of a cell body together with its prolongations, its axons and dendrites. The cell body varies from four to one hundred and fifty microns in diameter. The form of the cell varies according to the number, position and size of the dendrites. The existence of a cell wall is problematical. Several authors describe a cell wall of extreme tenuity, others deny its existence. The extension of the spongioplasm into the intercellular gray substance, described in the section on Relations of neurons, seems to be evidence against the presence of the cell wall. The peripheral prolongations of the sensory nerves and the axons of the motor nerves are alike called axis cylinders. The structure of the sensory fiber of the adult is not to be distinguished, histologically, from the motor axon. In the ordinary nerve trunk the two classes of fibers are intermingled with the fine, non-medullated sympathetic axons (fibers of Remak) to make what is called the mixed nerve. The nerve trunk is enclosed by the perineurium and is divided more or less completely into bundles by the endoneurium and the epineurium, of all which are connective tissue sheaths for the support of the nerve fibers and their vessels. The strength of the nerve fibers depends upon the toughness of these connective tissue sheaths. The coverings of the nerve fibers, together with the fibers themselves, are nourished as are other tissues, by blood vessels and lymphatics. These are subject to variations in size, according to changes in the systemic blood pressure, and they are innervated by vaso-motor nerves, as are most of the blood vessels of the body. Thus the nerve trunks are subject to hyperemic and ischemic conditions as a result of abnormal vaso-motor impulses, as are other tissues and organs. In addition to the etiological factors of neuralgia and neuritis which are already fairly well known, the place of such circulatory changes must be recognized. While the neuritis of a severe form is usually due to alcoholism, lead, or some other chronic poisoning, the milder types and the neuralgias are often caused by the same structural abnormalities found to be efficient causes of hyperemias and congestions in other tissues, i.e., slight malpositions of vertebrae or ribs, abnormally contracted muscles, reflex irritation from other organs of the body, including the various sources of peripheral irritation, and by the poisons resulting from the retention of the autogenic wastes, or from the use of drugs. Since the neurilemma is continuous with the sheaths of the sensory ganglia, and since these also are nourished by the blood vessels and lymphaticss, themselves innervated by vaso-motor nerves, it is evident that the same abnormal conditions may bring about pain and hyperesthesias due to the ganglionic condition and not associated with motor or vaso-motor disturbance. Such cases are very resistant to ordinary methods of treatment; drugs only purchase temporary relief at the expense of greater pain later. In such cases the correction of the structural abnormalities may permit the normal circulation to be established, and recovery must follow in the degree possible to tissues which have been more or less injured, not only by the original cause of the disturbance, but by the efforts to reduce the pain. The effects of separation from the nucleus are very well shown in the phenomena of Wallerian degeneration, so called after Waller, who gave the first exact description of the condition. When any nerve fiber is cut, the part which is separated from the cell body undergoes a series of changes whose sum is called by this name. The nerve fiber itself begins to show the change within a few hours after section. The fiber becomes granular in appearance, the granules increase in size, and undergo fatty degeneration. The medullary sheath becomes degenerated into droplets, at first very small but increasing in size. These structures are absorbed, probably in part by the lymphatics and veins, and in part by being used as food by surrounding tissues. The neurilemma cells, which are, of course, not injured except at the very point of section, begin to multiply very rapidly. This multiplication may be due in part to the presence of the increased food supply, but is more probably due to the irritation of the disintegrating nerve fiber and myelin. If regeneration does not occur, these neurilemma cells die and are themselves absorbed. The rapid multiplication of the neurilemma cells produces a solid cord of connective tissues of an embryonic type. The center of this cord, perhaps because of pressure, perhaps because of the lack of nutrition, contains no nuclei. This central portion is called the band fiber. Under normal conditions, following section of a nerve trunk, regeneration occurs. The conditions most favorable to regeneration are important. The ends of the injured nerve should be brought as closely together as possible, and should be sewed together. The structures to which the nerve is distributed must be kept in as nearly as possible a normal condition. This is best secured, in the case of the muscles, by massage and by electrical stimulation, applied with care. In the case of the skin, for the sake of sensation, the massage is most helpful. The general health of the patient must be kept as nearly perfect as possible, and his attention should be directed to the paralyzed part of the body. The descending impulses from the cerebrum seem to stimulate the cells in the cord which are in closest relation to the injured fibers by the attentive attitude and the efforts toward movements, and regeneration seems to be somewhat facilitated in this way. Regeneration occurs most readily and most completely in patients who are young and in good health. The process of regeneration follows the course which is to be expected from the physiological conditions. The process of degeneration affects the central end of the stump for a distance of one or several segments. At the upper limit of the area of degeneration the stump of the nerve fiber becomes swollen and bulbous. From this bulbous extremity several fine fibers shoot out, directed peripherally. These fibers are of variable size. One of them soon attains a certain superiority, and the others become atrophied and lost. The remaining fiber grows toward the periphery at a rate approximating a millimeter a day, penetrates the band fiber formed by the parenchymous cells of the multiplying neurilemma, and ultimately makes connection with the structures originally innervated by the injured nerve. In its essential features the process of regeneration of nerve fibers follows the same biological laws which govern the process of regeneration in all living structures. There is nothing beyond the normal conditions which can add to the completeness of the regeneration nor increase the rate of the growth. No drug or stimulant can be used advantageously, nothing can be done which is of the least benefit except to give the growing fibers the rest which they had during their period of most rapid growth, and to keep the muscles and the sensory structures in as nearly as possible their normal condition. |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}